Candida albicans is a ubiquitous commensal of the mammalian microbiome and the most prevalent fungal pathogen of humans. It can cause superficial infections of the oropharynx, vagina, skin, and nails. In susceptible patients, C. albicans can enter the bloodstream and cause a frequently fatal disseminated infection. Morphological plasticity is its defining feature and is critical for its pathogenesis. A cell-type transition between yeast and hyphal morphologies in C. albicans was thought to underlie much of the variation in virulence observed in different host tissues. In addition, opaque, gray and gastrointestinally induced transition (GUT) cell types were recently reported that exhibit marked differences in vitro and in animal models of commensalism and disease. In this review, we explore the characteristics of these cell types of C. albicans. We highlight emerging knowledge about the associations of these different morphotypes with different host niches and virulence potential, as well as the environmental cues and signalling pathways that are involved in the morphological transitions.
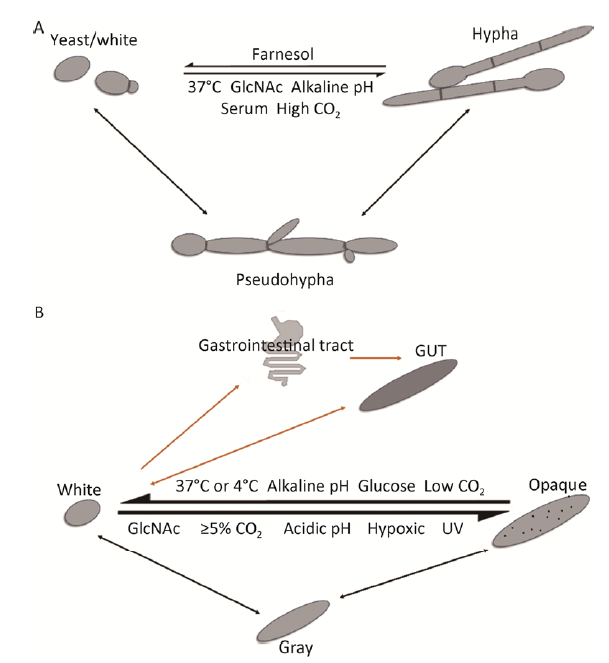
Fig. 1 Morphology and transformation of Candida albicans. A: Candida albicans’ yeast form cells (or white cells), pseudohyphae and hyphae can undergo reversible transformation under different environmental conditions; B: White-gray-opaque transformation (black arrow) and white-GUT transformation (orange arrow) of Candida albicans.
白念珠菌的酵母态、菌丝和假菌丝可以自身细胞类型稳定繁殖,也可以在形态发生过程中产生其他细胞类型。外界环境信号和内在的信号通路共同调控白念珠菌的形态转换,而这种形态转换与其致病性密切相关。白念珠菌酵母型细胞定植于黏膜或皮肤的表面,通常不引起宿主的免疫反应,但容易通过血液循环系统在宿主体内传播(Sudbery 2011);菌丝细胞在侵染宿主组织和穿透上皮细胞方面的能力较强(Phan et al. 2007),在逃逸巨噬细胞吞噬方面也具有重要的作用(Lorenz et al. 2004)。
1.2 白念珠菌white-opaque形态转换
白念珠菌除了能进行酵母-菌丝形态转换之外,还能进行white-opaque双稳态转换(图1B)。其最先是在一个白念珠菌临床分离菌WO-1中基于罕见的细胞和菌落形态的体外观察中发现的(Slutsky et al. 1987)。White细胞呈椭球至圆球形,在固体培养基上菌落呈奶油色,光亮、半圆形外观,其交配能力较opaque状态的白念珠菌弱;而opaque细胞在显微镜下是伸长的圆柱状,胞内通常含有大液泡,体积也较white细胞大约3倍,而且细胞表面有较小的“pimples”(通过扫描电子显微镜检测到的未知生物作用的突起)(Anderson et al. 1990),在固体培养基上形成较为粗糙的灰色扁平状菌落,其交配效率大约是white细胞的100万倍。White和opaque细胞的细胞壁通透性也不同,在添加红色染料荧光桃红B(phloxine B)的固体平板上,white细胞形成白色菌落,而opaque细胞则吸收染料形成红色或粉红色菌落(Anderson & Soll 1987)。菌丝的生长需要环境因子如37℃和血清等的持续刺激才能维持并且不能随着传代而遗传(Whiteway & Oberholzer 2004),而white和opaque两种形态细胞都可以稳定遗传并维持其原有形态几十代(Slutsky et al. 1987)。
最近有报道发现白念珠菌有一种类似于opaque细胞的gray细胞。Gray细胞形态可稳定遗传,并能够与white和opaque形态进行相互转换,形成一种“white-gray-opaque”三稳态转换系统(Tao et al. 2014)(图1B)。能够进行white-gray-opaque 形态转换的白念珠菌white状态的细胞在营养丰富的YEPD培养基上可以转换为gray细胞,但将其培养在营养缺乏的培养基(如Lee’s培养基)上,在加入N-乙酰葡糖胺(GlcNAc)和较高的CO2条件下会形成opaque细胞。Gray细胞呈较长的圆柱体形,但细胞体积较opaque和white细胞都小,且细胞表面没有“pimples”。在添加红色染料荧光桃红B的固体培养基上gray细胞形成浅红色或粉红色的菌落。Gray细胞能够进行交配,但其交配效率较低,介于white和opaque细胞之间。
在小鼠系统感染模型中,white细胞毒性最强,其次是opaque细胞,而gray细胞的毒性最弱。哺乳动物感染模型的初步研究表明opaque和gray细胞可能有对宿主上皮表面更高的适应能力(Xie et al. 2013;Tao et al. 2014),这说明白念珠菌面对不同的宿主环境,3种不同形态细胞表现不同的毒性特征。
1.4 白念珠菌的white-GUT形态转换
白念珠菌“GUT”细胞是在筛选感染小鼠消化道内共生状态的细胞时发现的(Pande et al. 2013)。将不同的白念珠菌突变株定植于小鼠胃肠道中,在该实验条件下宿主小鼠与高浓度的白念珠菌共生,仍保持健康状态。通过检测这些突变株在胃肠道共生的适应性,发现efg1突变体表现出增强的共生性;而wor1突变株在该模型中显示出明显降低的竞争性。有趣的是,Efg1和Wor1是一对在表达上相互抑制的转录因子。野生型白念珠菌在宿主消化道内传播时WOR1基因的表达量提高了10 000倍,而且通过异源的强启动子(WOR1OE)过表达WOR1,菌株表现出超强的共生竞争性,这些充分说明Wor1对白念珠菌共生适应性的促进作用。与opaque细胞相似,GUT细胞比同源white细胞形态更伸长,产生较暗的菌落,但是细胞表面没有“pimples”而且不能进行交配。研究表明,GUT细胞重新被引入到小鼠体内时,立刻表现出超强的共生竞争性,进一步证实GUT细胞比white和opaque细胞更适应胃肠道共生模型。综上这些数据表明,哺乳动物胃肠道的信号可能诱导白念珠菌表达WOR1并由white细胞转变为GUT细胞,而且这种GUT细胞需要持续性的胃肠道的信号诱导,否则会回复到white细胞。与white细胞相比,GUT细胞利用葡萄糖和吸收铁的能力出现下调,同时利用GlcNAc和短链脂肪酸的能力增强,因此GUT细胞似乎是在优化代谢以便其在哺乳动物消化道中作为共生体而生存(Tsong et al. 2003;Pande et al. 2013)(图1B)。
2.1.1 环境因子调控白念珠菌菌丝发生:人体的生理温度37℃是白念珠菌菌丝生长的最适温度(Whiteway & Oberholzer 2004)。血清和GlcNAc是白念珠菌菌丝生长的两种诱导因子。血清诱导菌丝生长的成分可能是肽聚糖,人体自身不能合成肽聚糖,血清中的这种成分可能主要来源于肠道共生细菌代谢(Xu et al. 2008),而且GlcNAc是消化道黏膜和细菌细胞壁的组成成分(Levitt & Bond 1970)。此外,高浓度CO2能够快速地诱导白念珠菌酵母形态向菌丝形态转换(Klengel et al. 2005),pH值在6.0-7.0时也有助于形成菌丝。细胞间的群体感应效应也会影响白念珠菌细胞的形态,高细胞密度时白念珠菌分泌的群体感应分子法尼醇(farnesol)对菌丝生长起明显的抑制作用,使得白念珠菌倾向于酵母形态的生长,而低细胞密度时更倾向于菌丝生长(Kumamoto 2005)。
White和opaque两种细胞形态形成菌丝的能力存在明显差异。在常规实验室培养条件下,opaque细胞不容易形成菌丝,很多能够诱导white细胞菌丝生长的环境因子对opaque细胞并不起作用(Ernst 2000)。但在某些特殊的环境条件下仍然可见opaque细胞菌丝生长现象,例如细胞贴壁生长时可以形成菌丝(Anderson et al. 1989)。低磷和山梨醇培养基可以特异性地诱导opaque细胞形成菌丝,而对white细胞的菌丝生长不起作用(Si et al. 2013)。
2.1.2 环境因子调控白念珠菌white-opaque形态转换:白念珠菌white和opaque表型之间的转换对环境条件高度敏感:GlcNAc、5%以上的CO2和酸性pH都有利于切换到opaque形态(Huang et al. 2009;Huang et al. 2010;Sun et al. 2015),而葡萄糖、低CO2水平、碱性pH和哺乳动物体温37℃促进回到white形态。White和opaque细胞在不同温度下表现出的稳定性也不同:室温条件下(22-25℃)opaque形态较稳定,而宿主生理温度(37℃)或低温(4℃)都不利于opaque细胞形态的维持(Rikkerink et al. 1988;Bergen et al. 1990)。研究发现,宿主皮肤温度较适中(约32℃)有利于白念珠菌维持opaque形态,因此皮肤可能是opaque形态较合适的定植部位(Soll et al. 2006)。CO2和GlcNAc均可在37℃时维持白念珠菌的opaque形态,并且在诱导white-opaque形态转换的过程中具有协同作用(Huang et al. 2010)。
环境pH变化影响Rim8、Rim13和Rim20等因子的活性,继而调控锌指结构转录因子Rim101活性。在暴露于碱性pH的酵母态白念珠菌细胞中,pH传感途径通过蛋白水解激活Rim101转录因子,从而促进菌丝特异性基因的表达和菌丝生长。激活态的Rim101调控pH感应基因PHR1和PHR2表达,PHR1和PHR2编码两个细胞表面糖苷酶,可能直接作为pH感应受体起作用,从而调控白念珠菌在不同pH环境中的菌丝生长(Muhlschlegel & Fonzi 1997;Fonzi 1999;Davis et al. 2000)。
2.2.2 转录水平调控白念珠菌white-gray-opaque转换:White-opaque转换与否的关键在于调控因子Wor1(white-opaque regulator 1)表达水平的高低(Huang et al. 2006;Srikantha et al. 2009)。白念珠菌第5号染色体上存在与酿酒酵母MAT(mating-type)基因座类似的MTL(mating-type like)基因座,包含了MTLa1、MTLa2、MTLα1及MTLα2这4个基因(Magee & Magee 2000)。MTL交配基因座不仅控制白念珠菌的交配,同时还控制white-opaque形态转换。自然界中绝大部分的白念珠菌是MTLa/α杂合型菌株,能合成Mtla1和Mtlα2形成Mtla1/α2异源复合物结合到WOR1的启动子区域抑制WOR1表达和opaque细胞形态的形成,最终抑制白念珠菌的交配(Zordan et al. 2006)。因此自然界中很少有能进行交配的白念珠菌,长期以来国际上也一直认为白念珠菌只能以二倍体状态存在,不进行减数分裂和交配(Dolan & Fields 1991)。随后才发现白念珠菌的交配效率与white-opaque形态转换密切相关,只有opaque形态细胞才能够进行高效率的交配(Park et al. 2013),而且这种交配也只能在MTL纯合的opaque细胞之间发生(Xie et al. 2013)。不过后来发现,白念珠菌杂合型MTLa/α的OFR1突变株的opaque形态也可以分别与MTLα/∆型和MTLa/∆型opaque细胞进行交配(Sun et al. 2016)。
转录因子Czf1和Wor2可正向调节white-opaque转换,而转录因子Efg1可抑制white-opaque转换,维持细胞处于white状态。Czf1、Wor2和Efg1 3个转录因子与Wor1共同组成一个反馈环调控白念珠菌white-opaque形态转换,Wor1处于这个调控网络的中心地位。Wor1不仅能够结合到自身启动子区域调控自身表达,还能够结合到EFG1、CZF1和WOR2的启动子区域,抑制EFG1表达,激活CZF1和WOR2表达。因此,当WOR1表达增加时会抑制EFG1的表达并促进opaque的状态,而WOR1表达下降时会引起对EFG1表达的去抑制作用(Srikantha et al. 2000;Vinces & Kumamoto 2007;Zordan et al. 2007;Ramirez-Zavala et al. 2008)。此外染色体修饰也可以影响white-opaque转换的效率,例如降低组蛋白去乙酰化水平能促进细胞从white状态转换为opaque状态(Morschhauser 2010)。
EFG1和WOR1基因单独敲除的菌株都可以形成gray形态,而同时敲除WOR1和EFG1后,细胞则保持在gray细胞形态,这说明Wor1和Efg1都不是gray细胞形成所必需的转录因子,但在white-gray-opaque三稳态形态转换的调控中可能具有协同作用(Tao et al. 2014)。HXK1(hexokinase 1)基因编码的GlcNAc激酶在GlcNAc代谢中催化GlcNAc-6-磷酸的形成,有研究发现在MTL杂合菌株中敲除HXK1可以促进gray和opaque细胞的形成,并且该突变株在3种形态之间可以进行相互转换,说明Hxk1参与调控白念珠菌white- gray-opaque三稳态形态转换(Cao et al. 2016)。
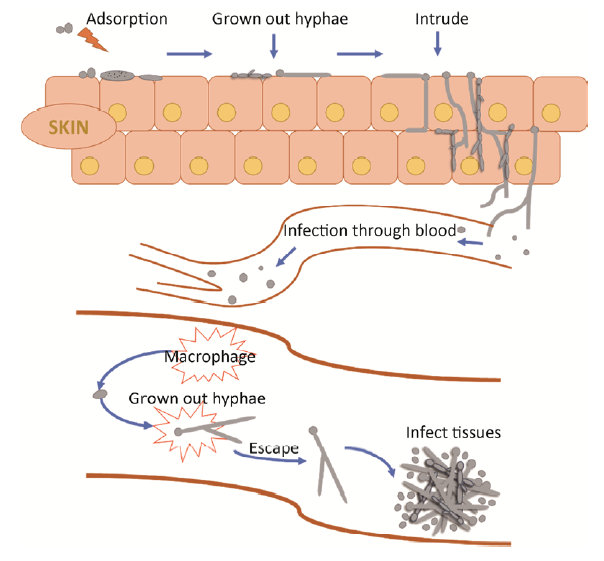
Fig. 2 Morphogenesis and pathogenicity of Candida albicans.
白念珠菌可表达黏附素以便其黏附到宿主的上皮细胞上。这个过程涉及白念珠菌的菌丝形成,因为菌丝更有利于其在组织表面黏附并为入侵做准备。菌丝细胞壁蛋白1(hyphal wall protein 1,Hwp1)是一种菌丝相关的糖基磷脂酰肌醇连接蛋白,参与菌丝细胞壁的构建。Hwp1是白念珠菌重要的黏附相关基因和菌丝相关基因,特异性地在白念珠菌菌丝中表达。有研究表明,Hwp1介导了白念珠菌对口腔黏膜上皮细胞的黏附,该基因的缺失可导致菌株致病力降低(Staab et al. 1999)。此外,凝集素样序列蛋白(agglutinin-like protein,ALS)家族基因是白念珠菌重要的黏附相关基因,对黏附和生物被膜的形成起着关键作用。其中,ALS3的缺失可造成菌株在多种环境中的黏附能力显著下降,其表达水平的高低直接影响了菌株的黏附能力和生物被膜形成能力(Chen & Lan 2015)。
随着白念珠菌形成的菌丝不断伸长,并入侵宿主组织,黏附、物理压力和真菌水解酶的分泌有助于菌丝入侵。分泌型天冬氨酸蛋白酶(secreted aspartyl protease,Sap)家族具有较高的蛋白水解酶活性,通过水解多种宿主底物(黏蛋白、角蛋白、胶原蛋白、E-钙黏着蛋白及宿主免疫应答相关的多种因子),促进白念珠菌汲取营养、附着和入侵宿主、损害宿主免疫应激反应等。其中,Sap4-Sap6在中性pH时的酵母态细胞向菌丝转变时表达,而唾液的pH值近中性,因此Sap4-Sap6可能在口腔念珠菌病的发病中起一定作用(Schaller et al. 2005)。白念珠菌入侵组织有2种途径,其中一种途径是被上皮细胞内吞。与酵母形态细胞相比,菌丝形态的白念珠菌更易被上皮细胞内吞。这可能是因为菌丝细胞壁中的凝集素样序列蛋白Als3结合到上皮细胞的E-钙黏素和N-钙黏素上从而诱导内吞(Phan et al. 2007)。另一种途径中白念珠菌形成的菌丝能有效地破坏上皮细胞而直接穿透组织。在感染过程中,白念珠菌的毒力与其菌丝形态密切相关,一些毒力因子的编码基因和菌丝特异性基因具有相似的表达调控机制(Gow & Hube 2012),在侵入组织的过程中白念珠菌菌丝发生常伴随着毒力因子如黏附素(Hwp1,Als3)和蛋白酶类(Sap4-Sap6)的表达。
白念珠菌的white-gray-opaque和white-GUT形态转换作为对宿主的适应机制与其致病性也有很大的关联。White细胞在系统感染中毒性较强,而由于opaque细胞能够分泌胞外天冬氨酸蛋白酶Sap降解宿主组织,故opaque细胞在皮肤和其他浅部感染中毒性比white细胞强(Kvaal et al. 1999)。而在含有牛血清白蛋白(BSA)的培养基中,gray细胞分泌Sap的能力比white和opaque细胞都强,在Lee’s葡萄糖培养条件下,opaque细胞的Sap活性比gray细胞强,说明白念珠菌面对不同的宿主环境,3种不同形态细胞可能表现出不同的毒性特征(Tao et al. 2014)。White和opaque细胞对宿主免疫细胞敏感性也不同,如white细胞更容易被巨噬细胞吞噬,而opaque细胞能有效地逃避宿主免疫细胞的识别(Lohse & Johnson 2008)。白念珠菌黏附到不同细胞表面需要不同种类的黏附素,有些黏附素在white细胞中高表达而有些黏附素在opaque细胞中高表达。有研究表明,white细胞更容易黏附在口腔上皮细胞,而opaque细胞更倾向于黏附在皮肤上,这就意味着white和opaque细胞对宿主的不同生态位具有不同的偏好性和致病性(Lohse & Johnson 2008)。GUT细胞作为一种在哺乳动物胃肠道中发现的白念珠菌新形态,其作为一种优势共生态提高了对宿主微环境的适应性,对白念珠菌的传播有很大作用(Pande et al. 2013)。
Candida albicans lacking the gene encoding the regulatory subunit of protein kinase A displays a defect in hyphal formation and an altered localization of the catalytic subunit
Candida albicans strains heterozygous and homozygous for mutations in mitogen-activated protein kinase signaling components have defects in hyphal development
.Proceedings of the National Academy of Sciences of the United States of America, 93(23): 13223-13228
Misexpression of the opaque-phase-specific gene PEP1 (SAP1) in the white phase of Candida albicans confers increased virulence in a mouse model of cutaneous infection.
White-opaque switching in natural MTLa/alpha isolates of Candida albicans: evolutionary implications for roles in host adaptation, pathogenesis, and sex.
Hypha formation in the white-opaque transition of Candida albicans.
1
1989
... White和opaque两种细胞形态形成菌丝的能力存在明显差异.在常规实验室培养条件下,opaque细胞不容易形成菌丝,很多能够诱导white细胞菌丝生长的环境因子对opaque细胞并不起作用(Ernst 2000).但在某些特殊的环境条件下仍然可见opaque细胞菌丝生长现象,例如细胞贴壁生长时可以形成菌丝(Anderson et al. 1989).低磷和山梨醇培养基可以特异性地诱导opaque细胞形成菌丝,而对white细胞的菌丝生长不起作用(Si et al. 2013). ...
Ultrastructure and antigenicity of the unique cell wall pimple of the Candida opaque phenotype.
1
1990
... 白念珠菌除了能进行酵母-菌丝形态转换之外,还能进行white-opaque双稳态转换(图1B).其最先是在一个白念珠菌临床分离菌WO-1中基于罕见的细胞和菌落形态的体外观察中发现的(Slutsky et al. 1987).White细胞呈椭球至圆球形,在固体培养基上菌落呈奶油色,光亮、半圆形外观,其交配能力较opaque状态的白念珠菌弱;而opaque细胞在显微镜下是伸长的圆柱状,胞内通常含有大液泡,体积也较white细胞大约3倍,而且细胞表面有较小的“pimples”(通过扫描电子显微镜检测到的未知生物作用的突起)(Anderson et al. 1990),在固体培养基上形成较为粗糙的灰色扁平状菌落,其交配效率大约是white细胞的100万倍.White和opaque细胞的细胞壁通透性也不同,在添加红色染料荧光桃红B(phloxine B)的固体平板上,white细胞形成白色菌落,而opaque细胞则吸收染料形成红色或粉红色菌落(Anderson & Soll 1987).菌丝的生长需要环境因子如37℃和血清等的持续刺激才能维持并且不能随着传代而遗传(Whiteway & Oberholzer 2004),而white和opaque两种形态细胞都可以稳定遗传并维持其原有形态几十代(Slutsky et al. 1987). ...
Unique phenotype of opaque cells in the white-opaque transition of Candida albicans.
1
1987
... 白念珠菌除了能进行酵母-菌丝形态转换之外,还能进行white-opaque双稳态转换(图1B).其最先是在一个白念珠菌临床分离菌WO-1中基于罕见的细胞和菌落形态的体外观察中发现的(Slutsky et al. 1987).White细胞呈椭球至圆球形,在固体培养基上菌落呈奶油色,光亮、半圆形外观,其交配能力较opaque状态的白念珠菌弱;而opaque细胞在显微镜下是伸长的圆柱状,胞内通常含有大液泡,体积也较white细胞大约3倍,而且细胞表面有较小的“pimples”(通过扫描电子显微镜检测到的未知生物作用的突起)(Anderson et al. 1990),在固体培养基上形成较为粗糙的灰色扁平状菌落,其交配效率大约是white细胞的100万倍.White和opaque细胞的细胞壁通透性也不同,在添加红色染料荧光桃红B(phloxine B)的固体平板上,white细胞形成白色菌落,而opaque细胞则吸收染料形成红色或粉红色菌落(Anderson & Soll 1987).菌丝的生长需要环境因子如37℃和血清等的持续刺激才能维持并且不能随着传代而遗传(Whiteway & Oberholzer 2004),而white和opaque两种形态细胞都可以稳定遗传并维持其原有形态几十代(Slutsky et al. 1987). ...
Switching at the cellular level in the white-opaque transition of Candida albicans.
1
1990
... 2.1.2 环境因子调控白念珠菌white-opaque形态转换:白念珠菌white和opaque表型之间的转换对环境条件高度敏感:GlcNAc、5%以上的CO2和酸性pH都有利于切换到opaque形态(Huang et al. 2009;Huang et al. 2010;Sun et al. 2015),而葡萄糖、低CO2水平、碱性pH和哺乳动物体温37℃促进回到white形态.White和opaque细胞在不同温度下表现出的稳定性也不同:室温条件下(22-25℃)opaque形态较稳定,而宿主生理温度(37℃)或低温(4℃)都不利于opaque细胞形态的维持(Rikkerink et al. 1988;Bergen et al. 1990).研究发现,宿主皮肤温度较适中(约32℃)有利于白念珠菌维持opaque形态,因此皮肤可能是opaque形态较合适的定植部位(Soll et al. 2006).CO2和GlcNAc均可在37℃时维持白念珠菌的opaque形态,并且在诱导white-opaque形态转换的过程中具有协同作用(Huang et al. 2010). ...
A potential phosphorylation site for an A-type kinase in the Efg1 regulator protein contributes to hyphal morphogenesis of Candida albicans.
1
2001
... 2.2.1 转录水平调控白念珠菌菌丝形成:cAMP依赖的蛋白激酶A(cAMP-dependent protein kinase A,cAMP-PKA)是诱导菌丝形成的主要通路(Sudbery 2011).cAMP-PKA信号通路是真核生物中保守的信号传导途径,在酿酒酵母和白念珠菌的形态转换过程中起关键的调控作用(Hogan & Sundstrom 2009),能激活促进菌丝特异性基因表达的转录因子(Bockmuhl et al. 2001).CO2、GlcNAc和血清等环境因子诱导的白念珠菌菌丝生长主要通过这条途径发挥作用(Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005).Ras1是保守的GTP酶(small GTPase)蛋白,位于cAMP/PKA通路的上游,结合GTP后具有活性,过表达活性形式的RAS1(RAS1 G13V)能够促进菌丝生长(Takai et al. 2001).CYR1(CDC35)是白念珠菌中唯一腺苷酸环化酶编码基因(Rocha et al. 2001).胞外信号分子通过Ras1激活Cyr1,催化ATP转化为cAMP,导致胞内cAMP水平升高.cAMP结合PKA复合体的调节亚基Bcy1,使其与催化亚基解离,从而激活PKA的催化亚基,磷酸化并激活下游相应的转录因子,调控白念珠菌的菌丝生长(Bockmuhl & Ernst 2001;Cassola et al. 2004).该信号通路最终激活转录因子Efg1和Flo8等,调控菌丝特异性基因(HWP1、ECE1等)的表达,进而调控菌丝生长(Zheng et al. 2004). ...
Distinct and redundant roles of the two protein kinase A isoforms Tpk1p and Tpk2p in morphogenesis and growth of Candida albicans.
1
2001
... 2.2.1 转录水平调控白念珠菌菌丝形成:cAMP依赖的蛋白激酶A(cAMP-dependent protein kinase A,cAMP-PKA)是诱导菌丝形成的主要通路(Sudbery 2011).cAMP-PKA信号通路是真核生物中保守的信号传导途径,在酿酒酵母和白念珠菌的形态转换过程中起关键的调控作用(Hogan & Sundstrom 2009),能激活促进菌丝特异性基因表达的转录因子(Bockmuhl et al. 2001).CO2、GlcNAc和血清等环境因子诱导的白念珠菌菌丝生长主要通过这条途径发挥作用(Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005).Ras1是保守的GTP酶(small GTPase)蛋白,位于cAMP/PKA通路的上游,结合GTP后具有活性,过表达活性形式的RAS1(RAS1 G13V)能够促进菌丝生长(Takai et al. 2001).CYR1(CDC35)是白念珠菌中唯一腺苷酸环化酶编码基因(Rocha et al. 2001).胞外信号分子通过Ras1激活Cyr1,催化ATP转化为cAMP,导致胞内cAMP水平升高.cAMP结合PKA复合体的调节亚基Bcy1,使其与催化亚基解离,从而激活PKA的催化亚基,磷酸化并激活下游相应的转录因子,调控白念珠菌的菌丝生长(Bockmuhl & Ernst 2001;Cassola et al. 2004).该信号通路最终激活转录因子Efg1和Flo8等,调控菌丝特异性基因(HWP1、ECE1等)的表达,进而调控菌丝生长(Zheng et al. 2004). ...
A characterization of pH-regulated dimorphism in Candida albicans.
1
1984
... 多种环境因子(如温度、血清、GlcNAc、低氧、CO2、pH、肽聚糖和氨基酸)都可通过信号转导通路组成的网络来调控白念珠菌的形态发生(Buffo et al. 1984;Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005;Xu et al. 2008). ...
Role of the N-acetylglucosamine kinase (Hxk1) in the regulation of white-gray-opaque tristable phenotypic transitions in C. albicans.
1
2016
... EFG1和WOR1基因单独敲除的菌株都可以形成gray形态,而同时敲除WOR1和EFG1后,细胞则保持在gray细胞形态,这说明Wor1和Efg1都不是gray细胞形成所必需的转录因子,但在white-gray-opaque三稳态形态转换的调控中可能具有协同作用(Tao et al. 2014).HXK1(hexokinase 1)基因编码的GlcNAc激酶在GlcNAc代谢中催化GlcNAc-6-磷酸的形成,有研究发现在MTL杂合菌株中敲除HXK1可以促进gray和opaque细胞的形成,并且该突变株在3种形态之间可以进行相互转换,说明Hxk1参与调控白念珠菌white- gray-opaque三稳态形态转换(Cao et al. 2016). ...
Candida albicans lacking the gene encoding the regulatory subunit of protein kinase A displays a defect in hyphal formation and an altered localization of the catalytic subunit
1
2004
... 2.2.1 转录水平调控白念珠菌菌丝形成:cAMP依赖的蛋白激酶A(cAMP-dependent protein kinase A,cAMP-PKA)是诱导菌丝形成的主要通路(Sudbery 2011).cAMP-PKA信号通路是真核生物中保守的信号传导途径,在酿酒酵母和白念珠菌的形态转换过程中起关键的调控作用(Hogan & Sundstrom 2009),能激活促进菌丝特异性基因表达的转录因子(Bockmuhl et al. 2001).CO2、GlcNAc和血清等环境因子诱导的白念珠菌菌丝生长主要通过这条途径发挥作用(Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005).Ras1是保守的GTP酶(small GTPase)蛋白,位于cAMP/PKA通路的上游,结合GTP后具有活性,过表达活性形式的RAS1(RAS1 G13V)能够促进菌丝生长(Takai et al. 2001).CYR1(CDC35)是白念珠菌中唯一腺苷酸环化酶编码基因(Rocha et al. 2001).胞外信号分子通过Ras1激活Cyr1,催化ATP转化为cAMP,导致胞内cAMP水平升高.cAMP结合PKA复合体的调节亚基Bcy1,使其与催化亚基解离,从而激活PKA的催化亚基,磷酸化并激活下游相应的转录因子,调控白念珠菌的菌丝生长(Bockmuhl & Ernst 2001;Cassola et al. 2004).该信号通路最终激活转录因子Efg1和Flo8等,调控菌丝特异性基因(HWP1、ECE1等)的表达,进而调控菌丝生长(Zheng et al. 2004). ...
Role of SFP1 in the regulation of Candida albicans biofilm formation.
1
2015
... 白念珠菌可表达黏附素以便其黏附到宿主的上皮细胞上.这个过程涉及白念珠菌的菌丝形成,因为菌丝更有利于其在组织表面黏附并为入侵做准备.菌丝细胞壁蛋白1(hyphal wall protein 1,Hwp1)是一种菌丝相关的糖基磷脂酰肌醇连接蛋白,参与菌丝细胞壁的构建.Hwp1是白念珠菌重要的黏附相关基因和菌丝相关基因,特异性地在白念珠菌菌丝中表达.有研究表明,Hwp1介导了白念珠菌对口腔黏膜上皮细胞的黏附,该基因的缺失可导致菌株致病力降低(Staab et al. 1999).此外,凝集素样序列蛋白(agglutinin-like protein,ALS)家族基因是白念珠菌重要的黏附相关基因,对黏附和生物被膜的形成起着关键作用.其中,ALS3的缺失可造成菌株在多种环境中的黏附能力显著下降,其表达水平的高低直接影响了菌株的黏附能力和生物被膜形成能力(Chen & Lan 2015). ...
Roles of the Candida albicans mitogen-activated protein kinase homolog, Cek1p, in hyphal development and systemic candidiasis.
1
1998
... CEK1-MAPK途径也可以促进菌丝发生:MAPK信号通路由3种连续激活的蛋白激酶[MAPKKK(Ste11)、MAPKK(Hst7)和MAPK(Cek1或Cek2)]组成,作用于下游的转录因子Cph1和Tec1等(Liu et al. 1994;Schweizer et al. 2000).酵母态白念珠菌在低氧或包埋培养条件下可以通过激活Cek1促进菌丝生长(Kohler & Fink 1996;Csank et al. 1998).阻断MAPK信号通路明显影响白念珠菌菌丝生长,但其影响仍不如阻断cAMP/PKA信号通路强.特别是在血清和GlcNAc等菌丝诱导因子作用下,MAPK信号通路相关突变株仍然能进行菌丝生长(Leberer et al. 1996). ...
RIM101-dependent and-independent pathways govern pH responses in Candida albicans.
1
2000
... 环境pH变化影响Rim8、Rim13和Rim20等因子的活性,继而调控锌指结构转录因子Rim101活性.在暴露于碱性pH的酵母态白念珠菌细胞中,pH传感途径通过蛋白水解激活Rim101转录因子,从而促进菌丝特异性基因的表达和菌丝生长.激活态的Rim101调控pH感应基因PHR1和PHR2表达,PHR1和PHR2编码两个细胞表面糖苷酶,可能直接作为pH感应受体起作用,从而调控白念珠菌在不同pH环境中的菌丝生长(Muhlschlegel & Fonzi 1997;Fonzi 1999;Davis et al. 2000). ...
Cell-type-specific transcription in yeast
1
1991
... 2.2.2 转录水平调控白念珠菌white-gray-opaque转换:White-opaque转换与否的关键在于调控因子Wor1(white-opaque regulator 1)表达水平的高低(Huang et al. 2006;Srikantha et al. 2009).白念珠菌第5号染色体上存在与酿酒酵母MAT(mating-type)基因座类似的MTL(mating-type like)基因座,包含了MTLa1、MTLa2、MTLα1及MTLα2这4个基因(Magee & Magee 2000).MTL交配基因座不仅控制白念珠菌的交配,同时还控制white-opaque形态转换.自然界中绝大部分的白念珠菌是MTLa/α杂合型菌株,能合成Mtla1和Mtlα2形成Mtla1/α2异源复合物结合到WOR1的启动子区域抑制WOR1表达和opaque细胞形态的形成,最终抑制白念珠菌的交配(Zordan et al. 2006).因此自然界中很少有能进行交配的白念珠菌,长期以来国际上也一直认为白念珠菌只能以二倍体状态存在,不进行减数分裂和交配(Dolan & Fields 1991).随后才发现白念珠菌的交配效率与white-opaque形态转换密切相关,只有opaque形态细胞才能够进行高效率的交配(Park et al. 2013),而且这种交配也只能在MTL纯合的opaque细胞之间发生(Xie et al. 2013).不过后来发现,白念珠菌杂合型MTLa/α的OFR1突变株的opaque形态也可以分别与MTLα/∆型和MTLa/∆型opaque细胞进行交配(Sun et al. 2016). ...
Transcription factors in Candida albicans - environmental control of morphogenesis.
1
2000
... White和opaque两种细胞形态形成菌丝的能力存在明显差异.在常规实验室培养条件下,opaque细胞不容易形成菌丝,很多能够诱导white细胞菌丝生长的环境因子对opaque细胞并不起作用(Ernst 2000).但在某些特殊的环境条件下仍然可见opaque细胞菌丝生长现象,例如细胞贴壁生长时可以形成菌丝(Anderson et al. 1989).低磷和山梨醇培养基可以特异性地诱导opaque细胞形成菌丝,而对white细胞的菌丝生长不起作用(Si et al. 2013). ...
Ras signaling is required for serum-induced hyphal differentiation in Candida albicans.
2
1999
... 多种环境因子(如温度、血清、GlcNAc、低氧、CO2、pH、肽聚糖和氨基酸)都可通过信号转导通路组成的网络来调控白念珠菌的形态发生(Buffo et al. 1984;Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005;Xu et al. 2008). ...
... 2.2.1 转录水平调控白念珠菌菌丝形成:cAMP依赖的蛋白激酶A(cAMP-dependent protein kinase A,cAMP-PKA)是诱导菌丝形成的主要通路(Sudbery 2011).cAMP-PKA信号通路是真核生物中保守的信号传导途径,在酿酒酵母和白念珠菌的形态转换过程中起关键的调控作用(Hogan & Sundstrom 2009),能激活促进菌丝特异性基因表达的转录因子(Bockmuhl et al. 2001).CO2、GlcNAc和血清等环境因子诱导的白念珠菌菌丝生长主要通过这条途径发挥作用(Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005).Ras1是保守的GTP酶(small GTPase)蛋白,位于cAMP/PKA通路的上游,结合GTP后具有活性,过表达活性形式的RAS1(RAS1 G13V)能够促进菌丝生长(Takai et al. 2001).CYR1(CDC35)是白念珠菌中唯一腺苷酸环化酶编码基因(Rocha et al. 2001).胞外信号分子通过Ras1激活Cyr1,催化ATP转化为cAMP,导致胞内cAMP水平升高.cAMP结合PKA复合体的调节亚基Bcy1,使其与催化亚基解离,从而激活PKA的催化亚基,磷酸化并激活下游相应的转录因子,调控白念珠菌的菌丝生长(Bockmuhl & Ernst 2001;Cassola et al. 2004).该信号通路最终激活转录因子Efg1和Flo8等,调控菌丝特异性基因(HWP1、ECE1等)的表达,进而调控菌丝生长(Zheng et al. 2004). ...
Topographic diversity of fungal and bacterial communities in human skin
1
2013
... 白念珠菌Candida albicans,又称白色念珠菌或白假丝酵母菌,是目前公认的人类最常见的真菌共生菌和病原体,存在于70%以上的正常人体的口腔、表皮、上呼吸道、胃肠道及阴道等处(Ghannoum et al. 2010;Findley et al. 2013;Hoffmann et al. 2013;Merenstein et al. 2013).在健康人体内,这种共生通常是良性的,一般不会引起疾病或仅引起轻微的浅表感染,但是当人体免疫系统受损或体内正常微生物菌群失衡时,该菌的过度生长会引起较严重的浅部感染(如鹅口疮和阴道炎),甚至可能导致致命性的深部器官或系统感染(如败血症)(Odds 1988).虽然目前有多种抗真菌治疗手段,但系统性念珠菌病仍有较高的致死率.目前,念珠菌属病原真菌所引起的侵袭性血液感染在美国医院中排名高达第3-4位(Pfaller & Diekema 2007).据估计,大约75%的女性一生中至少一次患有外阴阴道念珠菌病,40%-50%至少经历过一次感染(Hurley & De Louvois 1979). ...
PHR1 and PHR2 of Candida albicans encode putative glycosidases required for proper cross-linking of beta-1,3- and beta-1,6-glucans.
1
1999
... 环境pH变化影响Rim8、Rim13和Rim20等因子的活性,继而调控锌指结构转录因子Rim101活性.在暴露于碱性pH的酵母态白念珠菌细胞中,pH传感途径通过蛋白水解激活Rim101转录因子,从而促进菌丝特异性基因的表达和菌丝生长.激活态的Rim101调控pH感应基因PHR1和PHR2表达,PHR1和PHR2编码两个细胞表面糖苷酶,可能直接作为pH感应受体起作用,从而调控白念珠菌在不同pH环境中的菌丝生长(Muhlschlegel & Fonzi 1997;Fonzi 1999;Davis et al. 2000). ...
Characterization of the oral fungal microbiome (mycobiome) in healthy individuals
1
2010
... 白念珠菌Candida albicans,又称白色念珠菌或白假丝酵母菌,是目前公认的人类最常见的真菌共生菌和病原体,存在于70%以上的正常人体的口腔、表皮、上呼吸道、胃肠道及阴道等处(Ghannoum et al. 2010;Findley et al. 2013;Hoffmann et al. 2013;Merenstein et al. 2013).在健康人体内,这种共生通常是良性的,一般不会引起疾病或仅引起轻微的浅表感染,但是当人体免疫系统受损或体内正常微生物菌群失衡时,该菌的过度生长会引起较严重的浅部感染(如鹅口疮和阴道炎),甚至可能导致致命性的深部器官或系统感染(如败血症)(Odds 1988).虽然目前有多种抗真菌治疗手段,但系统性念珠菌病仍有较高的致死率.目前,念珠菌属病原真菌所引起的侵袭性血液感染在美国医院中排名高达第3-4位(Pfaller & Diekema 2007).据估计,大约75%的女性一生中至少一次患有外阴阴道念珠菌病,40%-50%至少经历过一次感染(Hurley & De Louvois 1979). ...
Importance of the Candida albicans cell wall during commensalism and infection.
1
2012
... 随着白念珠菌形成的菌丝不断伸长,并入侵宿主组织,黏附、物理压力和真菌水解酶的分泌有助于菌丝入侵.分泌型天冬氨酸蛋白酶(secreted aspartyl protease,Sap)家族具有较高的蛋白水解酶活性,通过水解多种宿主底物(黏蛋白、角蛋白、胶原蛋白、E-钙黏着蛋白及宿主免疫应答相关的多种因子),促进白念珠菌汲取营养、附着和入侵宿主、损害宿主免疫应激反应等.其中,Sap4-Sap6在中性pH时的酵母态细胞向菌丝转变时表达,而唾液的pH值近中性,因此Sap4-Sap6可能在口腔念珠菌病的发病中起一定作用(Schaller et al. 2005).白念珠菌入侵组织有2种途径,其中一种途径是被上皮细胞内吞.与酵母形态细胞相比,菌丝形态的白念珠菌更易被上皮细胞内吞.这可能是因为菌丝细胞壁中的凝集素样序列蛋白Als3结合到上皮细胞的E-钙黏素和N-钙黏素上从而诱导内吞(Phan et al. 2007).另一种途径中白念珠菌形成的菌丝能有效地破坏上皮细胞而直接穿透组织.在感染过程中,白念珠菌的毒力与其菌丝形态密切相关,一些毒力因子的编码基因和菌丝特异性基因具有相似的表达调控机制(Gow & Hube 2012),在侵入组织的过程中白念珠菌菌丝发生常伴随着毒力因子如黏附素(Hwp1,Als3)和蛋白酶类(Sap4-Sap6)的表达. ...
Archaea and fungi of the human gut microbiome: correlations with diet and bacterial residents
1
2013
... 白念珠菌Candida albicans,又称白色念珠菌或白假丝酵母菌,是目前公认的人类最常见的真菌共生菌和病原体,存在于70%以上的正常人体的口腔、表皮、上呼吸道、胃肠道及阴道等处(Ghannoum et al. 2010;Findley et al. 2013;Hoffmann et al. 2013;Merenstein et al. 2013).在健康人体内,这种共生通常是良性的,一般不会引起疾病或仅引起轻微的浅表感染,但是当人体免疫系统受损或体内正常微生物菌群失衡时,该菌的过度生长会引起较严重的浅部感染(如鹅口疮和阴道炎),甚至可能导致致命性的深部器官或系统感染(如败血症)(Odds 1988).虽然目前有多种抗真菌治疗手段,但系统性念珠菌病仍有较高的致死率.目前,念珠菌属病原真菌所引起的侵袭性血液感染在美国医院中排名高达第3-4位(Pfaller & Diekema 2007).据估计,大约75%的女性一生中至少一次患有外阴阴道念珠菌病,40%-50%至少经历过一次感染(Hurley & De Louvois 1979). ...
The Ras/cAMP/PKA signaling pathway and virulence in Candida albicans.
1
2009
... 2.2.1 转录水平调控白念珠菌菌丝形成:cAMP依赖的蛋白激酶A(cAMP-dependent protein kinase A,cAMP-PKA)是诱导菌丝形成的主要通路(Sudbery 2011).cAMP-PKA信号通路是真核生物中保守的信号传导途径,在酿酒酵母和白念珠菌的形态转换过程中起关键的调控作用(Hogan & Sundstrom 2009),能激活促进菌丝特异性基因表达的转录因子(Bockmuhl et al. 2001).CO2、GlcNAc和血清等环境因子诱导的白念珠菌菌丝生长主要通过这条途径发挥作用(Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005).Ras1是保守的GTP酶(small GTPase)蛋白,位于cAMP/PKA通路的上游,结合GTP后具有活性,过表达活性形式的RAS1(RAS1 G13V)能够促进菌丝生长(Takai et al. 2001).CYR1(CDC35)是白念珠菌中唯一腺苷酸环化酶编码基因(Rocha et al. 2001).胞外信号分子通过Ras1激活Cyr1,催化ATP转化为cAMP,导致胞内cAMP水平升高.cAMP结合PKA复合体的调节亚基Bcy1,使其与催化亚基解离,从而激活PKA的催化亚基,磷酸化并激活下游相应的转录因子,调控白念珠菌的菌丝生长(Bockmuhl & Ernst 2001;Cassola et al. 2004).该信号通路最终激活转录因子Efg1和Flo8等,调控菌丝特异性基因(HWP1、ECE1等)的表达,进而调控菌丝生长(Zheng et al. 2004). ...
Regulation of phenotypic transitions in the fungal pathogen Candida albicans.
1
2012
... 大部分真菌具有较强的形态可塑性,许多真菌在不同的生长环境以不同的形态存在.鉴于白念珠菌的整个生命周期发生在哺乳动物宿主中,而且具有特定风险的病人几乎所有器官都可能感染白念珠菌,所以为了适应宿主体内复杂的环境条件,白念珠菌进化出多种适应机制,其中白念珠菌形态发生与其环境适应能力以及致病性密切相关.在生长过程中,依据极性生长程度不同,白念珠菌可分为3种形态:酵母态、假菌丝和真菌丝.以及在某些特殊条件下形成的经典细胞类型——厚垣孢子(Jansons & Nickerson 1970).白念珠菌还可以在正常酵母形态(白菌,white)和伸长的细胞形态(灰菌,opaque)之间转换.Gray形态和GUT(gastrointestinally-induced transition,肠胃诱发的转变)形态是近年来在白念珠菌中发现的两种新形态,gray细胞可与white和opaque细胞形成white-gray-opaque三稳态转换系统(Tao et al. 2014);GUT细胞可与white细胞之间发生转换(Pande et al. 2013).白念珠菌不同形态的细胞在基因表达谱、毒性和交配能力等多方面具有明显差异,并与其致病性关系密切(Huang 2012). ...
CO(2) regulates white-to-opaque switching in Candida albicans.
1
2009
... 2.1.2 环境因子调控白念珠菌white-opaque形态转换:白念珠菌white和opaque表型之间的转换对环境条件高度敏感:GlcNAc、5%以上的CO2和酸性pH都有利于切换到opaque形态(Huang et al. 2009;Huang et al. 2010;Sun et al. 2015),而葡萄糖、低CO2水平、碱性pH和哺乳动物体温37℃促进回到white形态.White和opaque细胞在不同温度下表现出的稳定性也不同:室温条件下(22-25℃)opaque形态较稳定,而宿主生理温度(37℃)或低温(4℃)都不利于opaque细胞形态的维持(Rikkerink et al. 1988;Bergen et al. 1990).研究发现,宿主皮肤温度较适中(约32℃)有利于白念珠菌维持opaque形态,因此皮肤可能是opaque形态较合适的定植部位(Soll et al. 2006).CO2和GlcNAc均可在37℃时维持白念珠菌的opaque形态,并且在诱导white-opaque形态转换的过程中具有协同作用(Huang et al. 2010). ...
Bistable expression of WOR1, a master regulator of white-opaque switching in Candida albicans.
1
2006
... 2.2.2 转录水平调控白念珠菌white-gray-opaque转换:White-opaque转换与否的关键在于调控因子Wor1(white-opaque regulator 1)表达水平的高低(Huang et al. 2006;Srikantha et al. 2009).白念珠菌第5号染色体上存在与酿酒酵母MAT(mating-type)基因座类似的MTL(mating-type like)基因座,包含了MTLa1、MTLa2、MTLα1及MTLα2这4个基因(Magee & Magee 2000).MTL交配基因座不仅控制白念珠菌的交配,同时还控制white-opaque形态转换.自然界中绝大部分的白念珠菌是MTLa/α杂合型菌株,能合成Mtla1和Mtlα2形成Mtla1/α2异源复合物结合到WOR1的启动子区域抑制WOR1表达和opaque细胞形态的形成,最终抑制白念珠菌的交配(Zordan et al. 2006).因此自然界中很少有能进行交配的白念珠菌,长期以来国际上也一直认为白念珠菌只能以二倍体状态存在,不进行减数分裂和交配(Dolan & Fields 1991).随后才发现白念珠菌的交配效率与white-opaque形态转换密切相关,只有opaque形态细胞才能够进行高效率的交配(Park et al. 2013),而且这种交配也只能在MTL纯合的opaque细胞之间发生(Xie et al. 2013).不过后来发现,白念珠菌杂合型MTLa/α的OFR1突变株的opaque形态也可以分别与MTLα/∆型和MTLa/∆型opaque细胞进行交配(Sun et al. 2016). ...
N-acetylglucosamine induces white to opaque switching, a mating prerequisite in Candida albicans.
2
2010
... 2.1.2 环境因子调控白念珠菌white-opaque形态转换:白念珠菌white和opaque表型之间的转换对环境条件高度敏感:GlcNAc、5%以上的CO2和酸性pH都有利于切换到opaque形态(Huang et al. 2009;Huang et al. 2010;Sun et al. 2015),而葡萄糖、低CO2水平、碱性pH和哺乳动物体温37℃促进回到white形态.White和opaque细胞在不同温度下表现出的稳定性也不同:室温条件下(22-25℃)opaque形态较稳定,而宿主生理温度(37℃)或低温(4℃)都不利于opaque细胞形态的维持(Rikkerink et al. 1988;Bergen et al. 1990).研究发现,宿主皮肤温度较适中(约32℃)有利于白念珠菌维持opaque形态,因此皮肤可能是opaque形态较合适的定植部位(Soll et al. 2006).CO2和GlcNAc均可在37℃时维持白念珠菌的opaque形态,并且在诱导white-opaque形态转换的过程中具有协同作用(Huang et al. 2010). ...
... 和GlcNAc均可在37℃时维持白念珠菌的opaque形态,并且在诱导white-opaque形态转换的过程中具有协同作用(Huang et al. 2010). ...
Candida vaginitis
1
1979
... 白念珠菌Candida albicans,又称白色念珠菌或白假丝酵母菌,是目前公认的人类最常见的真菌共生菌和病原体,存在于70%以上的正常人体的口腔、表皮、上呼吸道、胃肠道及阴道等处(Ghannoum et al. 2010;Findley et al. 2013;Hoffmann et al. 2013;Merenstein et al. 2013).在健康人体内,这种共生通常是良性的,一般不会引起疾病或仅引起轻微的浅表感染,但是当人体免疫系统受损或体内正常微生物菌群失衡时,该菌的过度生长会引起较严重的浅部感染(如鹅口疮和阴道炎),甚至可能导致致命性的深部器官或系统感染(如败血症)(Odds 1988).虽然目前有多种抗真菌治疗手段,但系统性念珠菌病仍有较高的致死率.目前,念珠菌属病原真菌所引起的侵袭性血液感染在美国医院中排名高达第3-4位(Pfaller & Diekema 2007).据估计,大约75%的女性一生中至少一次患有外阴阴道念珠菌病,40%-50%至少经历过一次感染(Hurley & De Louvois 1979). ...
Induction, morphogenesis, and germination of the chlamydospore of Candida albicans.
1
1970
... 大部分真菌具有较强的形态可塑性,许多真菌在不同的生长环境以不同的形态存在.鉴于白念珠菌的整个生命周期发生在哺乳动物宿主中,而且具有特定风险的病人几乎所有器官都可能感染白念珠菌,所以为了适应宿主体内复杂的环境条件,白念珠菌进化出多种适应机制,其中白念珠菌形态发生与其环境适应能力以及致病性密切相关.在生长过程中,依据极性生长程度不同,白念珠菌可分为3种形态:酵母态、假菌丝和真菌丝.以及在某些特殊条件下形成的经典细胞类型——厚垣孢子(Jansons & Nickerson 1970).白念珠菌还可以在正常酵母形态(白菌,white)和伸长的细胞形态(灰菌,opaque)之间转换.Gray形态和GUT(gastrointestinally-induced transition,肠胃诱发的转变)形态是近年来在白念珠菌中发现的两种新形态,gray细胞可与white和opaque细胞形成white-gray-opaque三稳态转换系统(Tao et al. 2014);GUT细胞可与white细胞之间发生转换(Pande et al. 2013).白念珠菌不同形态的细胞在基因表达谱、毒性和交配能力等多方面具有明显差异,并与其致病性关系密切(Huang 2012). ...
Fungal adenylyl cyclase integrates CO2 sensing with cAMP signaling and virulence
3
2005
... 多种环境因子(如温度、血清、GlcNAc、低氧、CO2、pH、肽聚糖和氨基酸)都可通过信号转导通路组成的网络来调控白念珠菌的形态发生(Buffo et al. 1984;Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005;Xu et al. 2008). ...
... 2.1.1 环境因子调控白念珠菌菌丝发生:人体的生理温度37℃是白念珠菌菌丝生长的最适温度(Whiteway & Oberholzer 2004).血清和GlcNAc是白念珠菌菌丝生长的两种诱导因子.血清诱导菌丝生长的成分可能是肽聚糖,人体自身不能合成肽聚糖,血清中的这种成分可能主要来源于肠道共生细菌代谢(Xu et al. 2008),而且GlcNAc是消化道黏膜和细菌细胞壁的组成成分(Levitt & Bond 1970).此外,高浓度CO2能够快速地诱导白念珠菌酵母形态向菌丝形态转换(Klengel et al. 2005),pH值在6.0-7.0时也有助于形成菌丝.细胞间的群体感应效应也会影响白念珠菌细胞的形态,高细胞密度时白念珠菌分泌的群体感应分子法尼醇(farnesol)对菌丝生长起明显的抑制作用,使得白念珠菌倾向于酵母形态的生长,而低细胞密度时更倾向于菌丝生长(Kumamoto 2005). ...
... 2.2.1 转录水平调控白念珠菌菌丝形成:cAMP依赖的蛋白激酶A(cAMP-dependent protein kinase A,cAMP-PKA)是诱导菌丝形成的主要通路(Sudbery 2011).cAMP-PKA信号通路是真核生物中保守的信号传导途径,在酿酒酵母和白念珠菌的形态转换过程中起关键的调控作用(Hogan & Sundstrom 2009),能激活促进菌丝特异性基因表达的转录因子(Bockmuhl et al. 2001).CO2、GlcNAc和血清等环境因子诱导的白念珠菌菌丝生长主要通过这条途径发挥作用(Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005).Ras1是保守的GTP酶(small GTPase)蛋白,位于cAMP/PKA通路的上游,结合GTP后具有活性,过表达活性形式的RAS1(RAS1 G13V)能够促进菌丝生长(Takai et al. 2001).CYR1(CDC35)是白念珠菌中唯一腺苷酸环化酶编码基因(Rocha et al. 2001).胞外信号分子通过Ras1激活Cyr1,催化ATP转化为cAMP,导致胞内cAMP水平升高.cAMP结合PKA复合体的调节亚基Bcy1,使其与催化亚基解离,从而激活PKA的催化亚基,磷酸化并激活下游相应的转录因子,调控白念珠菌的菌丝生长(Bockmuhl & Ernst 2001;Cassola et al. 2004).该信号通路最终激活转录因子Efg1和Flo8等,调控菌丝特异性基因(HWP1、ECE1等)的表达,进而调控菌丝生长(Zheng et al. 2004). ...
Candida albicans strains heterozygous and homozygous for mutations in mitogen-activated protein kinase signaling components have defects in hyphal development
1
1996
... CEK1-MAPK途径也可以促进菌丝发生:MAPK信号通路由3种连续激活的蛋白激酶[MAPKKK(Ste11)、MAPKK(Hst7)和MAPK(Cek1或Cek2)]组成,作用于下游的转录因子Cph1和Tec1等(Liu et al. 1994;Schweizer et al. 2000).酵母态白念珠菌在低氧或包埋培养条件下可以通过激活Cek1促进菌丝生长(Kohler & Fink 1996;Csank et al. 1998).阻断MAPK信号通路明显影响白念珠菌菌丝生长,但其影响仍不如阻断cAMP/PKA信号通路强.特别是在血清和GlcNAc等菌丝诱导因子作用下,MAPK信号通路相关突变株仍然能进行菌丝生长(Leberer et al. 1996). ...
A contact-activated kinase signals Candida albicans invasive growth and biofilm development.
1
2005
... 2.1.1 环境因子调控白念珠菌菌丝发生:人体的生理温度37℃是白念珠菌菌丝生长的最适温度(Whiteway & Oberholzer 2004).血清和GlcNAc是白念珠菌菌丝生长的两种诱导因子.血清诱导菌丝生长的成分可能是肽聚糖,人体自身不能合成肽聚糖,血清中的这种成分可能主要来源于肠道共生细菌代谢(Xu et al. 2008),而且GlcNAc是消化道黏膜和细菌细胞壁的组成成分(Levitt & Bond 1970).此外,高浓度CO2能够快速地诱导白念珠菌酵母形态向菌丝形态转换(Klengel et al. 2005),pH值在6.0-7.0时也有助于形成菌丝.细胞间的群体感应效应也会影响白念珠菌细胞的形态,高细胞密度时白念珠菌分泌的群体感应分子法尼醇(farnesol)对菌丝生长起明显的抑制作用,使得白念珠菌倾向于酵母形态的生长,而低细胞密度时更倾向于菌丝生长(Kumamoto 2005). ...
Misexpression of the opaque-phase-specific gene PEP1 (SAP1) in the white phase of Candida albicans confers increased virulence in a mouse model of cutaneous infection.
1
1999
... 白念珠菌的white-gray-opaque和white-GUT形态转换作为对宿主的适应机制与其致病性也有很大的关联.White细胞在系统感染中毒性较强,而由于opaque细胞能够分泌胞外天冬氨酸蛋白酶Sap降解宿主组织,故opaque细胞在皮肤和其他浅部感染中毒性比white细胞强(Kvaal et al. 1999).而在含有牛血清白蛋白(BSA)的培养基中,gray细胞分泌Sap的能力比white和opaque细胞都强,在Lee’s葡萄糖培养条件下,opaque细胞的Sap活性比gray细胞强,说明白念珠菌面对不同的宿主环境,3种不同形态细胞可能表现出不同的毒性特征(Tao et al. 2014).White和opaque细胞对宿主免疫细胞敏感性也不同,如white细胞更容易被巨噬细胞吞噬,而opaque细胞能有效地逃避宿主免疫细胞的识别(Lohse & Johnson 2008).白念珠菌黏附到不同细胞表面需要不同种类的黏附素,有些黏附素在white细胞中高表达而有些黏附素在opaque细胞中高表达.有研究表明,white细胞更容易黏附在口腔上皮细胞,而opaque细胞更倾向于黏附在皮肤上,这就意味着white和opaque细胞对宿主的不同生态位具有不同的偏好性和致病性(Lohse & Johnson 2008).GUT细胞作为一种在哺乳动物胃肠道中发现的白念珠菌新形态,其作为一种优势共生态提高了对宿主微环境的适应性,对白念珠菌的传播有很大作用(Pande et al. 2013). ...
Metabolic specialization associated with phenotypic switching in Candida albicans.
1
2002
... 白念珠菌white和opaque细胞的基因组DNA序列完全相同,因此这种形态的转换主要受表观遗传的调控.White和opaque细胞各自表达一系列特异基因,有超过1 000个基因的表达发生变化,包括重要的交配和代谢基因(Lan et al. 2002;Tsong et al. 2003;Tuch et al. 2010).WH11、EFG1和发酵代谢相关基因在white细胞中特异性高表达,而OP4、SAP4和氧化代谢相关基因在opaque细胞中特异性表达(Morrow et al. 1993;Srikantha & Soll 1993). ...
Signal transduction through homologs of the Ste20p and Ste7p protein kinases can trigger hyphal formation in the pathogenic fungus Candida albicans.
1
1996
... CEK1-MAPK途径也可以促进菌丝发生:MAPK信号通路由3种连续激活的蛋白激酶[MAPKKK(Ste11)、MAPKK(Hst7)和MAPK(Cek1或Cek2)]组成,作用于下游的转录因子Cph1和Tec1等(Liu et al. 1994;Schweizer et al. 2000).酵母态白念珠菌在低氧或包埋培养条件下可以通过激活Cek1促进菌丝生长(Kohler & Fink 1996;Csank et al. 1998).阻断MAPK信号通路明显影响白念珠菌菌丝生长,但其影响仍不如阻断cAMP/PKA信号通路强.特别是在血清和GlcNAc等菌丝诱导因子作用下,MAPK信号通路相关突变株仍然能进行菌丝生长(Leberer et al. 1996). ...
Volume, composition, and source of intestinal gas
1
1970
... 2.1.1 环境因子调控白念珠菌菌丝发生:人体的生理温度37℃是白念珠菌菌丝生长的最适温度(Whiteway & Oberholzer 2004).血清和GlcNAc是白念珠菌菌丝生长的两种诱导因子.血清诱导菌丝生长的成分可能是肽聚糖,人体自身不能合成肽聚糖,血清中的这种成分可能主要来源于肠道共生细菌代谢(Xu et al. 2008),而且GlcNAc是消化道黏膜和细菌细胞壁的组成成分(Levitt & Bond 1970).此外,高浓度CO2能够快速地诱导白念珠菌酵母形态向菌丝形态转换(Klengel et al. 2005),pH值在6.0-7.0时也有助于形成菌丝.细胞间的群体感应效应也会影响白念珠菌细胞的形态,高细胞密度时白念珠菌分泌的群体感应分子法尼醇(farnesol)对菌丝生长起明显的抑制作用,使得白念珠菌倾向于酵母形态的生长,而低细胞密度时更倾向于菌丝生长(Kumamoto 2005). ...
Suppression of hyphal formation in Candida albicans by mutation of a STE12 homolog.
1
1994
... CEK1-MAPK途径也可以促进菌丝发生:MAPK信号通路由3种连续激活的蛋白激酶[MAPKKK(Ste11)、MAPKK(Hst7)和MAPK(Cek1或Cek2)]组成,作用于下游的转录因子Cph1和Tec1等(Liu et al. 1994;Schweizer et al. 2000).酵母态白念珠菌在低氧或包埋培养条件下可以通过激活Cek1促进菌丝生长(Kohler & Fink 1996;Csank et al. 1998).阻断MAPK信号通路明显影响白念珠菌菌丝生长,但其影响仍不如阻断cAMP/PKA信号通路强.特别是在血清和GlcNAc等菌丝诱导因子作用下,MAPK信号通路相关突变株仍然能进行菌丝生长(Leberer et al. 1996). ...
Differential phagocytosis of white versus opaque Candida albicans by Drosophila and mouse phagocytes.
2
2008
... 白念珠菌的white-gray-opaque和white-GUT形态转换作为对宿主的适应机制与其致病性也有很大的关联.White细胞在系统感染中毒性较强,而由于opaque细胞能够分泌胞外天冬氨酸蛋白酶Sap降解宿主组织,故opaque细胞在皮肤和其他浅部感染中毒性比white细胞强(Kvaal et al. 1999).而在含有牛血清白蛋白(BSA)的培养基中,gray细胞分泌Sap的能力比white和opaque细胞都强,在Lee’s葡萄糖培养条件下,opaque细胞的Sap活性比gray细胞强,说明白念珠菌面对不同的宿主环境,3种不同形态细胞可能表现出不同的毒性特征(Tao et al. 2014).White和opaque细胞对宿主免疫细胞敏感性也不同,如white细胞更容易被巨噬细胞吞噬,而opaque细胞能有效地逃避宿主免疫细胞的识别(Lohse & Johnson 2008).白念珠菌黏附到不同细胞表面需要不同种类的黏附素,有些黏附素在white细胞中高表达而有些黏附素在opaque细胞中高表达.有研究表明,white细胞更容易黏附在口腔上皮细胞,而opaque细胞更倾向于黏附在皮肤上,这就意味着white和opaque细胞对宿主的不同生态位具有不同的偏好性和致病性(Lohse & Johnson 2008).GUT细胞作为一种在哺乳动物胃肠道中发现的白念珠菌新形态,其作为一种优势共生态提高了对宿主微环境的适应性,对白念珠菌的传播有很大作用(Pande et al. 2013). ...
... ).白念珠菌黏附到不同细胞表面需要不同种类的黏附素,有些黏附素在white细胞中高表达而有些黏附素在opaque细胞中高表达.有研究表明,white细胞更容易黏附在口腔上皮细胞,而opaque细胞更倾向于黏附在皮肤上,这就意味着white和opaque细胞对宿主的不同生态位具有不同的偏好性和致病性(Lohse & Johnson 2008).GUT细胞作为一种在哺乳动物胃肠道中发现的白念珠菌新形态,其作为一种优势共生态提高了对宿主微环境的适应性,对白念珠菌的传播有很大作用(Pande et al. 2013). ...
Transcriptional response of Candida albicans upon internalization by macrophages.
1
2004
... 白念珠菌的酵母态、菌丝和假菌丝可以自身细胞类型稳定繁殖,也可以在形态发生过程中产生其他细胞类型.外界环境信号和内在的信号通路共同调控白念珠菌的形态转换,而这种形态转换与其致病性密切相关.白念珠菌酵母型细胞定植于黏膜或皮肤的表面,通常不引起宿主的免疫反应,但容易通过血液循环系统在宿主体内传播(Sudbery 2011);菌丝细胞在侵染宿主组织和穿透上皮细胞方面的能力较强(Phan et al. 2007),在逃逸巨噬细胞吞噬方面也具有重要的作用(Lorenz et al. 2004). ...
Induction of mating in Candida albicans by construction of MTLa and MTLalpha strains.
1
2000
... 2.2.2 转录水平调控白念珠菌white-gray-opaque转换:White-opaque转换与否的关键在于调控因子Wor1(white-opaque regulator 1)表达水平的高低(Huang et al. 2006;Srikantha et al. 2009).白念珠菌第5号染色体上存在与酿酒酵母MAT(mating-type)基因座类似的MTL(mating-type like)基因座,包含了MTLa1、MTLa2、MTLα1及MTLα2这4个基因(Magee & Magee 2000).MTL交配基因座不仅控制白念珠菌的交配,同时还控制white-opaque形态转换.自然界中绝大部分的白念珠菌是MTLa/α杂合型菌株,能合成Mtla1和Mtlα2形成Mtla1/α2异源复合物结合到WOR1的启动子区域抑制WOR1表达和opaque细胞形态的形成,最终抑制白念珠菌的交配(Zordan et al. 2006).因此自然界中很少有能进行交配的白念珠菌,长期以来国际上也一直认为白念珠菌只能以二倍体状态存在,不进行减数分裂和交配(Dolan & Fields 1991).随后才发现白念珠菌的交配效率与white-opaque形态转换密切相关,只有opaque形态细胞才能够进行高效率的交配(Park et al. 2013),而且这种交配也只能在MTL纯合的opaque细胞之间发生(Xie et al. 2013).不过后来发现,白念珠菌杂合型MTLa/α的OFR1突变株的opaque形态也可以分别与MTLα/∆型和MTLa/∆型opaque细胞进行交配(Sun et al. 2016). ...
Cell cycle dynamics and quorum sensing in Candida albicans chlamydospores are distinct from budding and hyphal growth.
1
2005
... 其中,酵母态和菌丝(图1A)是人们研究的 最多的两种形态,而对假菌丝和厚垣孢子的认识相对较少.正常的酵母态细胞也称为“white”细胞,具有类似于酿酒酵母的圆形到椭圆形的细胞形态,通过芽殖繁殖,核分裂发生在母细胞和子细胞之间的交界处.由于子代细胞在胞质分裂后与母代完全分离,故酵母态白念珠菌被认为是单细胞(Sudbery et al. 2004).相比之下,菌丝细胞是细长的管状细胞,核分裂发生在菌丝子细胞内,随后一个子核迁移到母细胞内.菌丝细胞在细胞分裂后仍保持牢固的连接,随着细胞分裂的重复产生多细胞、多分支的丝状结构,称为菌丝体.形态上真菌丝是完全平行延伸的管状菌丝,并且在细胞间隔处没有缢缩.假菌丝细胞则兼有酵母态和菌丝的特征,假菌丝是一串伸长的酵母态细胞,各个细胞之间没有分离,其细胞间有明显的分界,形成缢缩.假菌丝易形成分支,这样可能有利于其汲取营养.与酵母菌和菌丝不同的是,没有已知的体外条件能诱导出单一稳定的假菌丝菌落(Sudbery 2011).厚垣孢子是在某些严酷条件下(如饥饿和缺氧),在体外观察到的较大的球形厚壁细胞(Whiteway & Bachewich 2007).在厚垣孢子的形态发生过程中,细胞转变成丝状生长,然后形成伸长的胚柄细胞,从而产生厚垣孢子.核分裂发生在胚柄母细胞内,随后将子代细胞核迁移到新生的厚垣孢子中(Martin et al. 2005). ...
Colonization by Candida species of the oral and vaginal mucosa in HIV-infected and noninfected women.
1
2013
... 白念珠菌Candida albicans,又称白色念珠菌或白假丝酵母菌,是目前公认的人类最常见的真菌共生菌和病原体,存在于70%以上的正常人体的口腔、表皮、上呼吸道、胃肠道及阴道等处(Ghannoum et al. 2010;Findley et al. 2013;Hoffmann et al. 2013;Merenstein et al. 2013).在健康人体内,这种共生通常是良性的,一般不会引起疾病或仅引起轻微的浅表感染,但是当人体免疫系统受损或体内正常微生物菌群失衡时,该菌的过度生长会引起较严重的浅部感染(如鹅口疮和阴道炎),甚至可能导致致命性的深部器官或系统感染(如败血症)(Odds 1988).虽然目前有多种抗真菌治疗手段,但系统性念珠菌病仍有较高的致死率.目前,念珠菌属病原真菌所引起的侵袭性血液感染在美国医院中排名高达第3-4位(Pfaller & Diekema 2007).据估计,大约75%的女性一生中至少一次患有外阴阴道念珠菌病,40%-50%至少经历过一次感染(Hurley & De Louvois 1979). ...
Coordinate regulation of two opaque-phase-specific genes during white-opaque switching in Candida albicans.
1
1993
... 白念珠菌white和opaque细胞的基因组DNA序列完全相同,因此这种形态的转换主要受表观遗传的调控.White和opaque细胞各自表达一系列特异基因,有超过1 000个基因的表达发生变化,包括重要的交配和代谢基因(Lan et al. 2002;Tsong et al. 2003;Tuch et al. 2010).WH11、EFG1和发酵代谢相关基因在white细胞中特异性高表达,而OP4、SAP4和氧化代谢相关基因在opaque细胞中特异性表达(Morrow et al. 1993;Srikantha & Soll 1993). ...
Regulation of white-opaque switching in Candida albicans.
1
2010
... 转录因子Czf1和Wor2可正向调节white-opaque转换,而转录因子Efg1可抑制white-opaque转换,维持细胞处于white状态.Czf1、Wor2和Efg1 3个转录因子与Wor1共同组成一个反馈环调控白念珠菌white-opaque形态转换,Wor1处于这个调控网络的中心地位.Wor1不仅能够结合到自身启动子区域调控自身表达,还能够结合到EFG1、CZF1和WOR2的启动子区域,抑制EFG1表达,激活CZF1和WOR2表达.因此,当WOR1表达增加时会抑制EFG1的表达并促进opaque的状态,而WOR1表达下降时会引起对EFG1表达的去抑制作用(Srikantha et al. 2000;Vinces & Kumamoto 2007;Zordan et al. 2007;Ramirez-Zavala et al. 2008).此外染色体修饰也可以影响white-opaque转换的效率,例如降低组蛋白去乙酰化水平能促进细胞从white状态转换为opaque状态(Morschhauser 2010). ...
PHR2 of Candida albicans encodes a functional homolog of the pH-regulated gene PHR1 with an inverted pattern of pH-dependent expression.
1
1997
... 环境pH变化影响Rim8、Rim13和Rim20等因子的活性,继而调控锌指结构转录因子Rim101活性.在暴露于碱性pH的酵母态白念珠菌细胞中,pH传感途径通过蛋白水解激活Rim101转录因子,从而促进菌丝特异性基因的表达和菌丝生长.激活态的Rim101调控pH感应基因PHR1和PHR2表达,PHR1和PHR2编码两个细胞表面糖苷酶,可能直接作为pH感应受体起作用,从而调控白念珠菌在不同pH环境中的菌丝生长(Muhlschlegel & Fonzi 1997;Fonzi 1999;Davis et al. 2000). ...
Candida and candidosis: a review and bibliography. 2nd edition.
1
1988
... 白念珠菌Candida albicans,又称白色念珠菌或白假丝酵母菌,是目前公认的人类最常见的真菌共生菌和病原体,存在于70%以上的正常人体的口腔、表皮、上呼吸道、胃肠道及阴道等处(Ghannoum et al. 2010;Findley et al. 2013;Hoffmann et al. 2013;Merenstein et al. 2013).在健康人体内,这种共生通常是良性的,一般不会引起疾病或仅引起轻微的浅表感染,但是当人体免疫系统受损或体内正常微生物菌群失衡时,该菌的过度生长会引起较严重的浅部感染(如鹅口疮和阴道炎),甚至可能导致致命性的深部器官或系统感染(如败血症)(Odds 1988).虽然目前有多种抗真菌治疗手段,但系统性念珠菌病仍有较高的致死率.目前,念珠菌属病原真菌所引起的侵袭性血液感染在美国医院中排名高达第3-4位(Pfaller & Diekema 2007).据估计,大约75%的女性一生中至少一次患有外阴阴道念珠菌病,40%-50%至少经历过一次感染(Hurley & De Louvois 1979). ...
Passage through the mammalian gut triggers a phenotypic switch that promotes Candida albicans commensalism.
4
2013
... 大部分真菌具有较强的形态可塑性,许多真菌在不同的生长环境以不同的形态存在.鉴于白念珠菌的整个生命周期发生在哺乳动物宿主中,而且具有特定风险的病人几乎所有器官都可能感染白念珠菌,所以为了适应宿主体内复杂的环境条件,白念珠菌进化出多种适应机制,其中白念珠菌形态发生与其环境适应能力以及致病性密切相关.在生长过程中,依据极性生长程度不同,白念珠菌可分为3种形态:酵母态、假菌丝和真菌丝.以及在某些特殊条件下形成的经典细胞类型——厚垣孢子(Jansons & Nickerson 1970).白念珠菌还可以在正常酵母形态(白菌,white)和伸长的细胞形态(灰菌,opaque)之间转换.Gray形态和GUT(gastrointestinally-induced transition,肠胃诱发的转变)形态是近年来在白念珠菌中发现的两种新形态,gray细胞可与white和opaque细胞形成white-gray-opaque三稳态转换系统(Tao et al. 2014);GUT细胞可与white细胞之间发生转换(Pande et al. 2013).白念珠菌不同形态的细胞在基因表达谱、毒性和交配能力等多方面具有明显差异,并与其致病性关系密切(Huang 2012). ...
... 白念珠菌“GUT”细胞是在筛选感染小鼠消化道内共生状态的细胞时发现的(Pande et al. 2013).将不同的白念珠菌突变株定植于小鼠胃肠道中,在该实验条件下宿主小鼠与高浓度的白念珠菌共生,仍保持健康状态.通过检测这些突变株在胃肠道共生的适应性,发现efg1突变体表现出增强的共生性;而wor1突变株在该模型中显示出明显降低的竞争性.有趣的是,Efg1和Wor1是一对在表达上相互抑制的转录因子.野生型白念珠菌在宿主消化道内传播时WOR1基因的表达量提高了10 000倍,而且通过异源的强启动子(WOR1OE)过表达WOR1,菌株表现出超强的共生竞争性,这些充分说明Wor1对白念珠菌共生适应性的促进作用.与opaque细胞相似,GUT细胞比同源white细胞形态更伸长,产生较暗的菌落,但是细胞表面没有“pimples”而且不能进行交配.研究表明,GUT细胞重新被引入到小鼠体内时,立刻表现出超强的共生竞争性,进一步证实GUT细胞比white和opaque细胞更适应胃肠道共生模型.综上这些数据表明,哺乳动物胃肠道的信号可能诱导白念珠菌表达WOR1并由white细胞转变为GUT细胞,而且这种GUT细胞需要持续性的胃肠道的信号诱导,否则会回复到white细胞.与white细胞相比,GUT细胞利用葡萄糖和吸收铁的能力出现下调,同时利用GlcNAc和短链脂肪酸的能力增强,因此GUT细胞似乎是在优化代谢以便其在哺乳动物消化道中作为共生体而生存(Tsong et al. 2003;Pande et al. 2013)(图1B). ...
... ;Pande et al. 2013)(图1B). ...
... 白念珠菌的white-gray-opaque和white-GUT形态转换作为对宿主的适应机制与其致病性也有很大的关联.White细胞在系统感染中毒性较强,而由于opaque细胞能够分泌胞外天冬氨酸蛋白酶Sap降解宿主组织,故opaque细胞在皮肤和其他浅部感染中毒性比white细胞强(Kvaal et al. 1999).而在含有牛血清白蛋白(BSA)的培养基中,gray细胞分泌Sap的能力比white和opaque细胞都强,在Lee’s葡萄糖培养条件下,opaque细胞的Sap活性比gray细胞强,说明白念珠菌面对不同的宿主环境,3种不同形态细胞可能表现出不同的毒性特征(Tao et al. 2014).White和opaque细胞对宿主免疫细胞敏感性也不同,如white细胞更容易被巨噬细胞吞噬,而opaque细胞能有效地逃避宿主免疫细胞的识别(Lohse & Johnson 2008).白念珠菌黏附到不同细胞表面需要不同种类的黏附素,有些黏附素在white细胞中高表达而有些黏附素在opaque细胞中高表达.有研究表明,white细胞更容易黏附在口腔上皮细胞,而opaque细胞更倾向于黏附在皮肤上,这就意味着white和opaque细胞对宿主的不同生态位具有不同的偏好性和致病性(Lohse & Johnson 2008).GUT细胞作为一种在哺乳动物胃肠道中发现的白念珠菌新形态,其作为一种优势共生态提高了对宿主微环境的适应性,对白念珠菌的传播有很大作用(Pande et al. 2013). ...
Candida albicans forms a specialized “sexual” as well as “pathogenic” biofilm
1
2013
... 2.2.2 转录水平调控白念珠菌white-gray-opaque转换:White-opaque转换与否的关键在于调控因子Wor1(white-opaque regulator 1)表达水平的高低(Huang et al. 2006;Srikantha et al. 2009).白念珠菌第5号染色体上存在与酿酒酵母MAT(mating-type)基因座类似的MTL(mating-type like)基因座,包含了MTLa1、MTLa2、MTLα1及MTLα2这4个基因(Magee & Magee 2000).MTL交配基因座不仅控制白念珠菌的交配,同时还控制white-opaque形态转换.自然界中绝大部分的白念珠菌是MTLa/α杂合型菌株,能合成Mtla1和Mtlα2形成Mtla1/α2异源复合物结合到WOR1的启动子区域抑制WOR1表达和opaque细胞形态的形成,最终抑制白念珠菌的交配(Zordan et al. 2006).因此自然界中很少有能进行交配的白念珠菌,长期以来国际上也一直认为白念珠菌只能以二倍体状态存在,不进行减数分裂和交配(Dolan & Fields 1991).随后才发现白念珠菌的交配效率与white-opaque形态转换密切相关,只有opaque形态细胞才能够进行高效率的交配(Park et al. 2013),而且这种交配也只能在MTL纯合的opaque细胞之间发生(Xie et al. 2013).不过后来发现,白念珠菌杂合型MTLa/α的OFR1突变株的opaque形态也可以分别与MTLα/∆型和MTLa/∆型opaque细胞进行交配(Sun et al. 2016). ...
Epidemiology of invasive candidiasis: a persistent public health problem
1
2007
... 白念珠菌Candida albicans,又称白色念珠菌或白假丝酵母菌,是目前公认的人类最常见的真菌共生菌和病原体,存在于70%以上的正常人体的口腔、表皮、上呼吸道、胃肠道及阴道等处(Ghannoum et al. 2010;Findley et al. 2013;Hoffmann et al. 2013;Merenstein et al. 2013).在健康人体内,这种共生通常是良性的,一般不会引起疾病或仅引起轻微的浅表感染,但是当人体免疫系统受损或体内正常微生物菌群失衡时,该菌的过度生长会引起较严重的浅部感染(如鹅口疮和阴道炎),甚至可能导致致命性的深部器官或系统感染(如败血症)(Odds 1988).虽然目前有多种抗真菌治疗手段,但系统性念珠菌病仍有较高的致死率.目前,念珠菌属病原真菌所引起的侵袭性血液感染在美国医院中排名高达第3-4位(Pfaller & Diekema 2007).据估计,大约75%的女性一生中至少一次患有外阴阴道念珠菌病,40%-50%至少经历过一次感染(Hurley & De Louvois 1979). ...
Als3 is a Candida albicans invasin that binds to cadherins and induces endocytosis by host cells.
2
2007
... 白念珠菌的酵母态、菌丝和假菌丝可以自身细胞类型稳定繁殖,也可以在形态发生过程中产生其他细胞类型.外界环境信号和内在的信号通路共同调控白念珠菌的形态转换,而这种形态转换与其致病性密切相关.白念珠菌酵母型细胞定植于黏膜或皮肤的表面,通常不引起宿主的免疫反应,但容易通过血液循环系统在宿主体内传播(Sudbery 2011);菌丝细胞在侵染宿主组织和穿透上皮细胞方面的能力较强(Phan et al. 2007),在逃逸巨噬细胞吞噬方面也具有重要的作用(Lorenz et al. 2004). ...
... 随着白念珠菌形成的菌丝不断伸长,并入侵宿主组织,黏附、物理压力和真菌水解酶的分泌有助于菌丝入侵.分泌型天冬氨酸蛋白酶(secreted aspartyl protease,Sap)家族具有较高的蛋白水解酶活性,通过水解多种宿主底物(黏蛋白、角蛋白、胶原蛋白、E-钙黏着蛋白及宿主免疫应答相关的多种因子),促进白念珠菌汲取营养、附着和入侵宿主、损害宿主免疫应激反应等.其中,Sap4-Sap6在中性pH时的酵母态细胞向菌丝转变时表达,而唾液的pH值近中性,因此Sap4-Sap6可能在口腔念珠菌病的发病中起一定作用(Schaller et al. 2005).白念珠菌入侵组织有2种途径,其中一种途径是被上皮细胞内吞.与酵母形态细胞相比,菌丝形态的白念珠菌更易被上皮细胞内吞.这可能是因为菌丝细胞壁中的凝集素样序列蛋白Als3结合到上皮细胞的E-钙黏素和N-钙黏素上从而诱导内吞(Phan et al. 2007).另一种途径中白念珠菌形成的菌丝能有效地破坏上皮细胞而直接穿透组织.在感染过程中,白念珠菌的毒力与其菌丝形态密切相关,一些毒力因子的编码基因和菌丝特异性基因具有相似的表达调控机制(Gow & Hube 2012),在侵入组织的过程中白念珠菌菌丝发生常伴随着毒力因子如黏附素(Hwp1,Als3)和蛋白酶类(Sap4-Sap6)的表达. ...
Environmental induction of white-opaque switching in Candida albicans.
1
2008
... 转录因子Czf1和Wor2可正向调节white-opaque转换,而转录因子Efg1可抑制white-opaque转换,维持细胞处于white状态.Czf1、Wor2和Efg1 3个转录因子与Wor1共同组成一个反馈环调控白念珠菌white-opaque形态转换,Wor1处于这个调控网络的中心地位.Wor1不仅能够结合到自身启动子区域调控自身表达,还能够结合到EFG1、CZF1和WOR2的启动子区域,抑制EFG1表达,激活CZF1和WOR2表达.因此,当WOR1表达增加时会抑制EFG1的表达并促进opaque的状态,而WOR1表达下降时会引起对EFG1表达的去抑制作用(Srikantha et al. 2000;Vinces & Kumamoto 2007;Zordan et al. 2007;Ramirez-Zavala et al. 2008).此外染色体修饰也可以影响white-opaque转换的效率,例如降低组蛋白去乙酰化水平能促进细胞从white状态转换为opaque状态(Morschhauser 2010). ...
Opaque-white phenotype transition: a programmed morphological transition in Candida albicans.
1
1988
... 2.1.2 环境因子调控白念珠菌white-opaque形态转换:白念珠菌white和opaque表型之间的转换对环境条件高度敏感:GlcNAc、5%以上的CO2和酸性pH都有利于切换到opaque形态(Huang et al. 2009;Huang et al. 2010;Sun et al. 2015),而葡萄糖、低CO2水平、碱性pH和哺乳动物体温37℃促进回到white形态.White和opaque细胞在不同温度下表现出的稳定性也不同:室温条件下(22-25℃)opaque形态较稳定,而宿主生理温度(37℃)或低温(4℃)都不利于opaque细胞形态的维持(Rikkerink et al. 1988;Bergen et al. 1990).研究发现,宿主皮肤温度较适中(约32℃)有利于白念珠菌维持opaque形态,因此皮肤可能是opaque形态较合适的定植部位(Soll et al. 2006).CO2和GlcNAc均可在37℃时维持白念珠菌的opaque形态,并且在诱导white-opaque形态转换的过程中具有协同作用(Huang et al. 2010). ...
Signaling through adenylyl cyclase is essential for hyphal growth and virulence in the pathogenic fungus Candida albicans.
3
2001
... 多种环境因子(如温度、血清、GlcNAc、低氧、CO2、pH、肽聚糖和氨基酸)都可通过信号转导通路组成的网络来调控白念珠菌的形态发生(Buffo et al. 1984;Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005;Xu et al. 2008). ...
... 2.2.1 转录水平调控白念珠菌菌丝形成:cAMP依赖的蛋白激酶A(cAMP-dependent protein kinase A,cAMP-PKA)是诱导菌丝形成的主要通路(Sudbery 2011).cAMP-PKA信号通路是真核生物中保守的信号传导途径,在酿酒酵母和白念珠菌的形态转换过程中起关键的调控作用(Hogan & Sundstrom 2009),能激活促进菌丝特异性基因表达的转录因子(Bockmuhl et al. 2001).CO2、GlcNAc和血清等环境因子诱导的白念珠菌菌丝生长主要通过这条途径发挥作用(Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005).Ras1是保守的GTP酶(small GTPase)蛋白,位于cAMP/PKA通路的上游,结合GTP后具有活性,过表达活性形式的RAS1(RAS1 G13V)能够促进菌丝生长(Takai et al. 2001).CYR1(CDC35)是白念珠菌中唯一腺苷酸环化酶编码基因(Rocha et al. 2001).胞外信号分子通过Ras1激活Cyr1,催化ATP转化为cAMP,导致胞内cAMP水平升高.cAMP结合PKA复合体的调节亚基Bcy1,使其与催化亚基解离,从而激活PKA的催化亚基,磷酸化并激活下游相应的转录因子,调控白念珠菌的菌丝生长(Bockmuhl & Ernst 2001;Cassola et al. 2004).该信号通路最终激活转录因子Efg1和Flo8等,调控菌丝特异性基因(HWP1、ECE1等)的表达,进而调控菌丝生长(Zheng et al. 2004). ...
... )是白念珠菌中唯一腺苷酸环化酶编码基因(Rocha et al. 2001).胞外信号分子通过Ras1激活Cyr1,催化ATP转化为cAMP,导致胞内cAMP水平升高.cAMP结合PKA复合体的调节亚基Bcy1,使其与催化亚基解离,从而激活PKA的催化亚基,磷酸化并激活下游相应的转录因子,调控白念珠菌的菌丝生长(Bockmuhl & Ernst 2001;Cassola et al. 2004).该信号通路最终激活转录因子Efg1和Flo8等,调控菌丝特异性基因(HWP1、ECE1等)的表达,进而调控菌丝生长(Zheng et al. 2004). ...
Hydrolytic enzymes as virulence factors of Candida albicans.
1
2005
... 随着白念珠菌形成的菌丝不断伸长,并入侵宿主组织,黏附、物理压力和真菌水解酶的分泌有助于菌丝入侵.分泌型天冬氨酸蛋白酶(secreted aspartyl protease,Sap)家族具有较高的蛋白水解酶活性,通过水解多种宿主底物(黏蛋白、角蛋白、胶原蛋白、E-钙黏着蛋白及宿主免疫应答相关的多种因子),促进白念珠菌汲取营养、附着和入侵宿主、损害宿主免疫应激反应等.其中,Sap4-Sap6在中性pH时的酵母态细胞向菌丝转变时表达,而唾液的pH值近中性,因此Sap4-Sap6可能在口腔念珠菌病的发病中起一定作用(Schaller et al. 2005).白念珠菌入侵组织有2种途径,其中一种途径是被上皮细胞内吞.与酵母形态细胞相比,菌丝形态的白念珠菌更易被上皮细胞内吞.这可能是因为菌丝细胞壁中的凝集素样序列蛋白Als3结合到上皮细胞的E-钙黏素和N-钙黏素上从而诱导内吞(Phan et al. 2007).另一种途径中白念珠菌形成的菌丝能有效地破坏上皮细胞而直接穿透组织.在感染过程中,白念珠菌的毒力与其菌丝形态密切相关,一些毒力因子的编码基因和菌丝特异性基因具有相似的表达调控机制(Gow & Hube 2012),在侵入组织的过程中白念珠菌菌丝发生常伴随着毒力因子如黏附素(Hwp1,Als3)和蛋白酶类(Sap4-Sap6)的表达. ...
The TEA/ATTS transcription factor CaTec1p regulates hyphal development and virulence in Candida albicans.
1
2000
... CEK1-MAPK途径也可以促进菌丝发生:MAPK信号通路由3种连续激活的蛋白激酶[MAPKKK(Ste11)、MAPKK(Hst7)和MAPK(Cek1或Cek2)]组成,作用于下游的转录因子Cph1和Tec1等(Liu et al. 1994;Schweizer et al. 2000).酵母态白念珠菌在低氧或包埋培养条件下可以通过激活Cek1促进菌丝生长(Kohler & Fink 1996;Csank et al. 1998).阻断MAPK信号通路明显影响白念珠菌菌丝生长,但其影响仍不如阻断cAMP/PKA信号通路强.特别是在血清和GlcNAc等菌丝诱导因子作用下,MAPK信号通路相关突变株仍然能进行菌丝生长(Leberer et al. 1996). ...
Candida albicans white and opaque cells undergo distinct programs of filamentous growth
1
2013
... White和opaque两种细胞形态形成菌丝的能力存在明显差异.在常规实验室培养条件下,opaque细胞不容易形成菌丝,很多能够诱导white细胞菌丝生长的环境因子对opaque细胞并不起作用(Ernst 2000).但在某些特殊的环境条件下仍然可见opaque细胞菌丝生长现象,例如细胞贴壁生长时可以形成菌丝(Anderson et al. 1989).低磷和山梨醇培养基可以特异性地诱导opaque细胞形成菌丝,而对white细胞的菌丝生长不起作用(Si et al. 2013). ...
“White-opaque transition”: a second high-frequency switching system in Candida albicans.
2
1987
... 白念珠菌除了能进行酵母-菌丝形态转换之外,还能进行white-opaque双稳态转换(图1B).其最先是在一个白念珠菌临床分离菌WO-1中基于罕见的细胞和菌落形态的体外观察中发现的(Slutsky et al. 1987).White细胞呈椭球至圆球形,在固体培养基上菌落呈奶油色,光亮、半圆形外观,其交配能力较opaque状态的白念珠菌弱;而opaque细胞在显微镜下是伸长的圆柱状,胞内通常含有大液泡,体积也较white细胞大约3倍,而且细胞表面有较小的“pimples”(通过扫描电子显微镜检测到的未知生物作用的突起)(Anderson et al. 1990),在固体培养基上形成较为粗糙的灰色扁平状菌落,其交配效率大约是white细胞的100万倍.White和opaque细胞的细胞壁通透性也不同,在添加红色染料荧光桃红B(phloxine B)的固体平板上,white细胞形成白色菌落,而opaque细胞则吸收染料形成红色或粉红色菌落(Anderson & Soll 1987).菌丝的生长需要环境因子如37℃和血清等的持续刺激才能维持并且不能随着传代而遗传(Whiteway & Oberholzer 2004),而white和opaque两种形态细胞都可以稳定遗传并维持其原有形态几十代(Slutsky et al. 1987). ...
... ),而white和opaque两种形态细胞都可以稳定遗传并维持其原有形态几十代(Slutsky et al. 1987). ...
The mating-type locus and mating of Candida albicans and Candida glabrata.
1
2006
... 2.1.2 环境因子调控白念珠菌white-opaque形态转换:白念珠菌white和opaque表型之间的转换对环境条件高度敏感:GlcNAc、5%以上的CO2和酸性pH都有利于切换到opaque形态(Huang et al. 2009;Huang et al. 2010;Sun et al. 2015),而葡萄糖、低CO2水平、碱性pH和哺乳动物体温37℃促进回到white形态.White和opaque细胞在不同温度下表现出的稳定性也不同:室温条件下(22-25℃)opaque形态较稳定,而宿主生理温度(37℃)或低温(4℃)都不利于opaque细胞形态的维持(Rikkerink et al. 1988;Bergen et al. 1990).研究发现,宿主皮肤温度较适中(约32℃)有利于白念珠菌维持opaque形态,因此皮肤可能是opaque形态较合适的定植部位(Soll et al. 2006).CO2和GlcNAc均可在37℃时维持白念珠菌的opaque形态,并且在诱导white-opaque形态转换的过程中具有协同作用(Huang et al. 2010). ...
TOS9 regulates white-opaque switching in Candida albicans.
1
2009
... 2.2.2 转录水平调控白念珠菌white-gray-opaque转换:White-opaque转换与否的关键在于调控因子Wor1(white-opaque regulator 1)表达水平的高低(Huang et al. 2006;Srikantha et al. 2009).白念珠菌第5号染色体上存在与酿酒酵母MAT(mating-type)基因座类似的MTL(mating-type like)基因座,包含了MTLa1、MTLa2、MTLα1及MTLα2这4个基因(Magee & Magee 2000).MTL交配基因座不仅控制白念珠菌的交配,同时还控制white-opaque形态转换.自然界中绝大部分的白念珠菌是MTLa/α杂合型菌株,能合成Mtla1和Mtlα2形成Mtla1/α2异源复合物结合到WOR1的启动子区域抑制WOR1表达和opaque细胞形态的形成,最终抑制白念珠菌的交配(Zordan et al. 2006).因此自然界中很少有能进行交配的白念珠菌,长期以来国际上也一直认为白念珠菌只能以二倍体状态存在,不进行减数分裂和交配(Dolan & Fields 1991).随后才发现白念珠菌的交配效率与white-opaque形态转换密切相关,只有opaque形态细胞才能够进行高效率的交配(Park et al. 2013),而且这种交配也只能在MTL纯合的opaque细胞之间发生(Xie et al. 2013).不过后来发现,白念珠菌杂合型MTLa/α的OFR1突变株的opaque形态也可以分别与MTLα/∆型和MTLa/∆型opaque细胞进行交配(Sun et al. 2016). ...
A white-specific gene in the white-opaque switching system of Candida albicans.
1
1993
... 白念珠菌white和opaque细胞的基因组DNA序列完全相同,因此这种形态的转换主要受表观遗传的调控.White和opaque细胞各自表达一系列特异基因,有超过1 000个基因的表达发生变化,包括重要的交配和代谢基因(Lan et al. 2002;Tsong et al. 2003;Tuch et al. 2010).WH11、EFG1和发酵代谢相关基因在white细胞中特异性高表达,而OP4、SAP4和氧化代谢相关基因在opaque细胞中特异性表达(Morrow et al. 1993;Srikantha & Soll 1993). ...
EFG1 null mutants of Candida albicans switch but cannot express the complete phenotype of white-phase budding cells.
1
2000
... 转录因子Czf1和Wor2可正向调节white-opaque转换,而转录因子Efg1可抑制white-opaque转换,维持细胞处于white状态.Czf1、Wor2和Efg1 3个转录因子与Wor1共同组成一个反馈环调控白念珠菌white-opaque形态转换,Wor1处于这个调控网络的中心地位.Wor1不仅能够结合到自身启动子区域调控自身表达,还能够结合到EFG1、CZF1和WOR2的启动子区域,抑制EFG1表达,激活CZF1和WOR2表达.因此,当WOR1表达增加时会抑制EFG1的表达并促进opaque的状态,而WOR1表达下降时会引起对EFG1表达的去抑制作用(Srikantha et al. 2000;Vinces & Kumamoto 2007;Zordan et al. 2007;Ramirez-Zavala et al. 2008).此外染色体修饰也可以影响white-opaque转换的效率,例如降低组蛋白去乙酰化水平能促进细胞从white状态转换为opaque状态(Morschhauser 2010). ...
Adhesive and mammalian transglutaminase substrate properties of Candida albicans Hwp1.
1
1999
... 白念珠菌可表达黏附素以便其黏附到宿主的上皮细胞上.这个过程涉及白念珠菌的菌丝形成,因为菌丝更有利于其在组织表面黏附并为入侵做准备.菌丝细胞壁蛋白1(hyphal wall protein 1,Hwp1)是一种菌丝相关的糖基磷脂酰肌醇连接蛋白,参与菌丝细胞壁的构建.Hwp1是白念珠菌重要的黏附相关基因和菌丝相关基因,特异性地在白念珠菌菌丝中表达.有研究表明,Hwp1介导了白念珠菌对口腔黏膜上皮细胞的黏附,该基因的缺失可导致菌株致病力降低(Staab et al. 1999).此外,凝集素样序列蛋白(agglutinin-like protein,ALS)家族基因是白念珠菌重要的黏附相关基因,对黏附和生物被膜的形成起着关键作用.其中,ALS3的缺失可造成菌株在多种环境中的黏附能力显著下降,其表达水平的高低直接影响了菌株的黏附能力和生物被膜形成能力(Chen & Lan 2015). ...
The distinct morphogenic states of Candida albicans.
1
2004
... 其中,酵母态和菌丝(图1A)是人们研究的 最多的两种形态,而对假菌丝和厚垣孢子的认识相对较少.正常的酵母态细胞也称为“white”细胞,具有类似于酿酒酵母的圆形到椭圆形的细胞形态,通过芽殖繁殖,核分裂发生在母细胞和子细胞之间的交界处.由于子代细胞在胞质分裂后与母代完全分离,故酵母态白念珠菌被认为是单细胞(Sudbery et al. 2004).相比之下,菌丝细胞是细长的管状细胞,核分裂发生在菌丝子细胞内,随后一个子核迁移到母细胞内.菌丝细胞在细胞分裂后仍保持牢固的连接,随着细胞分裂的重复产生多细胞、多分支的丝状结构,称为菌丝体.形态上真菌丝是完全平行延伸的管状菌丝,并且在细胞间隔处没有缢缩.假菌丝细胞则兼有酵母态和菌丝的特征,假菌丝是一串伸长的酵母态细胞,各个细胞之间没有分离,其细胞间有明显的分界,形成缢缩.假菌丝易形成分支,这样可能有利于其汲取营养.与酵母菌和菌丝不同的是,没有已知的体外条件能诱导出单一稳定的假菌丝菌落(Sudbery 2011).厚垣孢子是在某些严酷条件下(如饥饿和缺氧),在体外观察到的较大的球形厚壁细胞(Whiteway & Bachewich 2007).在厚垣孢子的形态发生过程中,细胞转变成丝状生长,然后形成伸长的胚柄细胞,从而产生厚垣孢子.核分裂发生在胚柄母细胞内,随后将子代细胞核迁移到新生的厚垣孢子中(Martin et al. 2005). ...
Growth of Candida albicans hyphae.
3
2011
... 其中,酵母态和菌丝(图1A)是人们研究的 最多的两种形态,而对假菌丝和厚垣孢子的认识相对较少.正常的酵母态细胞也称为“white”细胞,具有类似于酿酒酵母的圆形到椭圆形的细胞形态,通过芽殖繁殖,核分裂发生在母细胞和子细胞之间的交界处.由于子代细胞在胞质分裂后与母代完全分离,故酵母态白念珠菌被认为是单细胞(Sudbery et al. 2004).相比之下,菌丝细胞是细长的管状细胞,核分裂发生在菌丝子细胞内,随后一个子核迁移到母细胞内.菌丝细胞在细胞分裂后仍保持牢固的连接,随着细胞分裂的重复产生多细胞、多分支的丝状结构,称为菌丝体.形态上真菌丝是完全平行延伸的管状菌丝,并且在细胞间隔处没有缢缩.假菌丝细胞则兼有酵母态和菌丝的特征,假菌丝是一串伸长的酵母态细胞,各个细胞之间没有分离,其细胞间有明显的分界,形成缢缩.假菌丝易形成分支,这样可能有利于其汲取营养.与酵母菌和菌丝不同的是,没有已知的体外条件能诱导出单一稳定的假菌丝菌落(Sudbery 2011).厚垣孢子是在某些严酷条件下(如饥饿和缺氧),在体外观察到的较大的球形厚壁细胞(Whiteway & Bachewich 2007).在厚垣孢子的形态发生过程中,细胞转变成丝状生长,然后形成伸长的胚柄细胞,从而产生厚垣孢子.核分裂发生在胚柄母细胞内,随后将子代细胞核迁移到新生的厚垣孢子中(Martin et al. 2005). ...
... 白念珠菌的酵母态、菌丝和假菌丝可以自身细胞类型稳定繁殖,也可以在形态发生过程中产生其他细胞类型.外界环境信号和内在的信号通路共同调控白念珠菌的形态转换,而这种形态转换与其致病性密切相关.白念珠菌酵母型细胞定植于黏膜或皮肤的表面,通常不引起宿主的免疫反应,但容易通过血液循环系统在宿主体内传播(Sudbery 2011);菌丝细胞在侵染宿主组织和穿透上皮细胞方面的能力较强(Phan et al. 2007),在逃逸巨噬细胞吞噬方面也具有重要的作用(Lorenz et al. 2004). ...
... 2.2.1 转录水平调控白念珠菌菌丝形成:cAMP依赖的蛋白激酶A(cAMP-dependent protein kinase A,cAMP-PKA)是诱导菌丝形成的主要通路(Sudbery 2011).cAMP-PKA信号通路是真核生物中保守的信号传导途径,在酿酒酵母和白念珠菌的形态转换过程中起关键的调控作用(Hogan & Sundstrom 2009),能激活促进菌丝特异性基因表达的转录因子(Bockmuhl et al. 2001).CO2、GlcNAc和血清等环境因子诱导的白念珠菌菌丝生长主要通过这条途径发挥作用(Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005).Ras1是保守的GTP酶(small GTPase)蛋白,位于cAMP/PKA通路的上游,结合GTP后具有活性,过表达活性形式的RAS1(RAS1 G13V)能够促进菌丝生长(Takai et al. 2001).CYR1(CDC35)是白念珠菌中唯一腺苷酸环化酶编码基因(Rocha et al. 2001).胞外信号分子通过Ras1激活Cyr1,催化ATP转化为cAMP,导致胞内cAMP水平升高.cAMP结合PKA复合体的调节亚基Bcy1,使其与催化亚基解离,从而激活PKA的催化亚基,磷酸化并激活下游相应的转录因子,调控白念珠菌的菌丝生长(Bockmuhl & Ernst 2001;Cassola et al. 2004).该信号通路最终激活转录因子Efg1和Flo8等,调控菌丝特异性基因(HWP1、ECE1等)的表达,进而调控菌丝生长(Zheng et al. 2004). ...
pH Regulates White-opaque switching and sexual mating in Candida albicans.
1
2015
... 2.1.2 环境因子调控白念珠菌white-opaque形态转换:白念珠菌white和opaque表型之间的转换对环境条件高度敏感:GlcNAc、5%以上的CO2和酸性pH都有利于切换到opaque形态(Huang et al. 2009;Huang et al. 2010;Sun et al. 2015),而葡萄糖、低CO2水平、碱性pH和哺乳动物体温37℃促进回到white形态.White和opaque细胞在不同温度下表现出的稳定性也不同:室温条件下(22-25℃)opaque形态较稳定,而宿主生理温度(37℃)或低温(4℃)都不利于opaque细胞形态的维持(Rikkerink et al. 1988;Bergen et al. 1990).研究发现,宿主皮肤温度较适中(约32℃)有利于白念珠菌维持opaque形态,因此皮肤可能是opaque形态较合适的定植部位(Soll et al. 2006).CO2和GlcNAc均可在37℃时维持白念珠菌的opaque形态,并且在诱导white-opaque形态转换的过程中具有协同作用(Huang et al. 2010). ...
Deletion of a Yci1 domain protein of Candida albicans allows homothallic mating in MTL heterozygous cells.
1
2016
... 2.2.2 转录水平调控白念珠菌white-gray-opaque转换:White-opaque转换与否的关键在于调控因子Wor1(white-opaque regulator 1)表达水平的高低(Huang et al. 2006;Srikantha et al. 2009).白念珠菌第5号染色体上存在与酿酒酵母MAT(mating-type)基因座类似的MTL(mating-type like)基因座,包含了MTLa1、MTLa2、MTLα1及MTLα2这4个基因(Magee & Magee 2000).MTL交配基因座不仅控制白念珠菌的交配,同时还控制white-opaque形态转换.自然界中绝大部分的白念珠菌是MTLa/α杂合型菌株,能合成Mtla1和Mtlα2形成Mtla1/α2异源复合物结合到WOR1的启动子区域抑制WOR1表达和opaque细胞形态的形成,最终抑制白念珠菌的交配(Zordan et al. 2006).因此自然界中很少有能进行交配的白念珠菌,长期以来国际上也一直认为白念珠菌只能以二倍体状态存在,不进行减数分裂和交配(Dolan & Fields 1991).随后才发现白念珠菌的交配效率与white-opaque形态转换密切相关,只有opaque形态细胞才能够进行高效率的交配(Park et al. 2013),而且这种交配也只能在MTL纯合的opaque细胞之间发生(Xie et al. 2013).不过后来发现,白念珠菌杂合型MTLa/α的OFR1突变株的opaque形态也可以分别与MTLα/∆型和MTLa/∆型opaque细胞进行交配(Sun et al. 2016). ...
Small GTP-binding proteins
1
2001
... 2.2.1 转录水平调控白念珠菌菌丝形成:cAMP依赖的蛋白激酶A(cAMP-dependent protein kinase A,cAMP-PKA)是诱导菌丝形成的主要通路(Sudbery 2011).cAMP-PKA信号通路是真核生物中保守的信号传导途径,在酿酒酵母和白念珠菌的形态转换过程中起关键的调控作用(Hogan & Sundstrom 2009),能激活促进菌丝特异性基因表达的转录因子(Bockmuhl et al. 2001).CO2、GlcNAc和血清等环境因子诱导的白念珠菌菌丝生长主要通过这条途径发挥作用(Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005).Ras1是保守的GTP酶(small GTPase)蛋白,位于cAMP/PKA通路的上游,结合GTP后具有活性,过表达活性形式的RAS1(RAS1 G13V)能够促进菌丝生长(Takai et al. 2001).CYR1(CDC35)是白念珠菌中唯一腺苷酸环化酶编码基因(Rocha et al. 2001).胞外信号分子通过Ras1激活Cyr1,催化ATP转化为cAMP,导致胞内cAMP水平升高.cAMP结合PKA复合体的调节亚基Bcy1,使其与催化亚基解离,从而激活PKA的催化亚基,磷酸化并激活下游相应的转录因子,调控白念珠菌的菌丝生长(Bockmuhl & Ernst 2001;Cassola et al. 2004).该信号通路最终激活转录因子Efg1和Flo8等,调控菌丝特异性基因(HWP1、ECE1等)的表达,进而调控菌丝生长(Zheng et al. 2004). ...
Discovery of a “white-gray- opaque” tristable phenotypic switching system in Candida albicans: roles of non-genetic diversity in host adaptation.
5
2014
... 大部分真菌具有较强的形态可塑性,许多真菌在不同的生长环境以不同的形态存在.鉴于白念珠菌的整个生命周期发生在哺乳动物宿主中,而且具有特定风险的病人几乎所有器官都可能感染白念珠菌,所以为了适应宿主体内复杂的环境条件,白念珠菌进化出多种适应机制,其中白念珠菌形态发生与其环境适应能力以及致病性密切相关.在生长过程中,依据极性生长程度不同,白念珠菌可分为3种形态:酵母态、假菌丝和真菌丝.以及在某些特殊条件下形成的经典细胞类型——厚垣孢子(Jansons & Nickerson 1970).白念珠菌还可以在正常酵母形态(白菌,white)和伸长的细胞形态(灰菌,opaque)之间转换.Gray形态和GUT(gastrointestinally-induced transition,肠胃诱发的转变)形态是近年来在白念珠菌中发现的两种新形态,gray细胞可与white和opaque细胞形成white-gray-opaque三稳态转换系统(Tao et al. 2014);GUT细胞可与white细胞之间发生转换(Pande et al. 2013).白念珠菌不同形态的细胞在基因表达谱、毒性和交配能力等多方面具有明显差异,并与其致病性关系密切(Huang 2012). ...
... 最近有报道发现白念珠菌有一种类似于opaque细胞的gray细胞.Gray细胞形态可稳定遗传,并能够与white和opaque形态进行相互转换,形成一种“white-gray-opaque”三稳态转换系统(Tao et al. 2014)(图1B).能够进行white-gray-opaque 形态转换的白念珠菌white状态的细胞在营养丰富的YEPD培养基上可以转换为gray细胞,但将其培养在营养缺乏的培养基(如Lee’s培养基)上,在加入N-乙酰葡糖胺(GlcNAc)和较高的CO2条件下会形成opaque细胞.Gray细胞呈较长的圆柱体形,但细胞体积较opaque和white细胞都小,且细胞表面没有“pimples”.在添加红色染料荧光桃红B的固体培养基上gray细胞形成浅红色或粉红色的菌落.Gray细胞能够进行交配,但其交配效率较低,介于white和opaque细胞之间. ...
... 在小鼠系统感染模型中,white细胞毒性最强,其次是opaque细胞,而gray细胞的毒性最弱.哺乳动物感染模型的初步研究表明opaque和gray细胞可能有对宿主上皮表面更高的适应能力(Xie et al. 2013;Tao et al. 2014),这说明白念珠菌面对不同的宿主环境,3种不同形态细胞表现不同的毒性特征. ...
... EFG1和WOR1基因单独敲除的菌株都可以形成gray形态,而同时敲除WOR1和EFG1后,细胞则保持在gray细胞形态,这说明Wor1和Efg1都不是gray细胞形成所必需的转录因子,但在white-gray-opaque三稳态形态转换的调控中可能具有协同作用(Tao et al. 2014).HXK1(hexokinase 1)基因编码的GlcNAc激酶在GlcNAc代谢中催化GlcNAc-6-磷酸的形成,有研究发现在MTL杂合菌株中敲除HXK1可以促进gray和opaque细胞的形成,并且该突变株在3种形态之间可以进行相互转换,说明Hxk1参与调控白念珠菌white- gray-opaque三稳态形态转换(Cao et al. 2016). ...
... 白念珠菌的white-gray-opaque和white-GUT形态转换作为对宿主的适应机制与其致病性也有很大的关联.White细胞在系统感染中毒性较强,而由于opaque细胞能够分泌胞外天冬氨酸蛋白酶Sap降解宿主组织,故opaque细胞在皮肤和其他浅部感染中毒性比white细胞强(Kvaal et al. 1999).而在含有牛血清白蛋白(BSA)的培养基中,gray细胞分泌Sap的能力比white和opaque细胞都强,在Lee’s葡萄糖培养条件下,opaque细胞的Sap活性比gray细胞强,说明白念珠菌面对不同的宿主环境,3种不同形态细胞可能表现出不同的毒性特征(Tao et al. 2014).White和opaque细胞对宿主免疫细胞敏感性也不同,如white细胞更容易被巨噬细胞吞噬,而opaque细胞能有效地逃避宿主免疫细胞的识别(Lohse & Johnson 2008).白念珠菌黏附到不同细胞表面需要不同种类的黏附素,有些黏附素在white细胞中高表达而有些黏附素在opaque细胞中高表达.有研究表明,white细胞更容易黏附在口腔上皮细胞,而opaque细胞更倾向于黏附在皮肤上,这就意味着white和opaque细胞对宿主的不同生态位具有不同的偏好性和致病性(Lohse & Johnson 2008).GUT细胞作为一种在哺乳动物胃肠道中发现的白念珠菌新形态,其作为一种优势共生态提高了对宿主微环境的适应性,对白念珠菌的传播有很大作用(Pande et al. 2013). ...
Evolution of a combinatorial transcriptional circuit: a case study in yeasts
2
2003
... 白念珠菌white和opaque细胞的基因组DNA序列完全相同,因此这种形态的转换主要受表观遗传的调控.White和opaque细胞各自表达一系列特异基因,有超过1 000个基因的表达发生变化,包括重要的交配和代谢基因(Lan et al. 2002;Tsong et al. 2003;Tuch et al. 2010).WH11、EFG1和发酵代谢相关基因在white细胞中特异性高表达,而OP4、SAP4和氧化代谢相关基因在opaque细胞中特异性表达(Morrow et al. 1993;Srikantha & Soll 1993). ...
... 白念珠菌“GUT”细胞是在筛选感染小鼠消化道内共生状态的细胞时发现的(Pande et al. 2013).将不同的白念珠菌突变株定植于小鼠胃肠道中,在该实验条件下宿主小鼠与高浓度的白念珠菌共生,仍保持健康状态.通过检测这些突变株在胃肠道共生的适应性,发现efg1突变体表现出增强的共生性;而wor1突变株在该模型中显示出明显降低的竞争性.有趣的是,Efg1和Wor1是一对在表达上相互抑制的转录因子.野生型白念珠菌在宿主消化道内传播时WOR1基因的表达量提高了10 000倍,而且通过异源的强启动子(WOR1OE)过表达WOR1,菌株表现出超强的共生竞争性,这些充分说明Wor1对白念珠菌共生适应性的促进作用.与opaque细胞相似,GUT细胞比同源white细胞形态更伸长,产生较暗的菌落,但是细胞表面没有“pimples”而且不能进行交配.研究表明,GUT细胞重新被引入到小鼠体内时,立刻表现出超强的共生竞争性,进一步证实GUT细胞比white和opaque细胞更适应胃肠道共生模型.综上这些数据表明,哺乳动物胃肠道的信号可能诱导白念珠菌表达WOR1并由white细胞转变为GUT细胞,而且这种GUT细胞需要持续性的胃肠道的信号诱导,否则会回复到white细胞.与white细胞相比,GUT细胞利用葡萄糖和吸收铁的能力出现下调,同时利用GlcNAc和短链脂肪酸的能力增强,因此GUT细胞似乎是在优化代谢以便其在哺乳动物消化道中作为共生体而生存(Tsong et al. 2003;Pande et al. 2013)(图1B). ...
The transcriptomes of two heritable cell types illuminate the circuit governing their differentiation
1
2010
... 白念珠菌white和opaque细胞的基因组DNA序列完全相同,因此这种形态的转换主要受表观遗传的调控.White和opaque细胞各自表达一系列特异基因,有超过1 000个基因的表达发生变化,包括重要的交配和代谢基因(Lan et al. 2002;Tsong et al. 2003;Tuch et al. 2010).WH11、EFG1和发酵代谢相关基因在white细胞中特异性高表达,而OP4、SAP4和氧化代谢相关基因在opaque细胞中特异性表达(Morrow et al. 1993;Srikantha & Soll 1993). ...
The morphogenetic regulator Czf1p is a DNA-binding protein that regulates white opaque switching in Candida albicans.
1
2007
... 转录因子Czf1和Wor2可正向调节white-opaque转换,而转录因子Efg1可抑制white-opaque转换,维持细胞处于white状态.Czf1、Wor2和Efg1 3个转录因子与Wor1共同组成一个反馈环调控白念珠菌white-opaque形态转换,Wor1处于这个调控网络的中心地位.Wor1不仅能够结合到自身启动子区域调控自身表达,还能够结合到EFG1、CZF1和WOR2的启动子区域,抑制EFG1表达,激活CZF1和WOR2表达.因此,当WOR1表达增加时会抑制EFG1的表达并促进opaque的状态,而WOR1表达下降时会引起对EFG1表达的去抑制作用(Srikantha et al. 2000;Vinces & Kumamoto 2007;Zordan et al. 2007;Ramirez-Zavala et al. 2008).此外染色体修饰也可以影响white-opaque转换的效率,例如降低组蛋白去乙酰化水平能促进细胞从white状态转换为opaque状态(Morschhauser 2010). ...
Morphogenesis in Candida albicans.
1
2007
... 其中,酵母态和菌丝(图1A)是人们研究的 最多的两种形态,而对假菌丝和厚垣孢子的认识相对较少.正常的酵母态细胞也称为“white”细胞,具有类似于酿酒酵母的圆形到椭圆形的细胞形态,通过芽殖繁殖,核分裂发生在母细胞和子细胞之间的交界处.由于子代细胞在胞质分裂后与母代完全分离,故酵母态白念珠菌被认为是单细胞(Sudbery et al. 2004).相比之下,菌丝细胞是细长的管状细胞,核分裂发生在菌丝子细胞内,随后一个子核迁移到母细胞内.菌丝细胞在细胞分裂后仍保持牢固的连接,随着细胞分裂的重复产生多细胞、多分支的丝状结构,称为菌丝体.形态上真菌丝是完全平行延伸的管状菌丝,并且在细胞间隔处没有缢缩.假菌丝细胞则兼有酵母态和菌丝的特征,假菌丝是一串伸长的酵母态细胞,各个细胞之间没有分离,其细胞间有明显的分界,形成缢缩.假菌丝易形成分支,这样可能有利于其汲取营养.与酵母菌和菌丝不同的是,没有已知的体外条件能诱导出单一稳定的假菌丝菌落(Sudbery 2011).厚垣孢子是在某些严酷条件下(如饥饿和缺氧),在体外观察到的较大的球形厚壁细胞(Whiteway & Bachewich 2007).在厚垣孢子的形态发生过程中,细胞转变成丝状生长,然后形成伸长的胚柄细胞,从而产生厚垣孢子.核分裂发生在胚柄母细胞内,随后将子代细胞核迁移到新生的厚垣孢子中(Martin et al. 2005). ...
Candida morphogenesis and host-pathogen interactions
2
2004
... 白念珠菌除了能进行酵母-菌丝形态转换之外,还能进行white-opaque双稳态转换(图1B).其最先是在一个白念珠菌临床分离菌WO-1中基于罕见的细胞和菌落形态的体外观察中发现的(Slutsky et al. 1987).White细胞呈椭球至圆球形,在固体培养基上菌落呈奶油色,光亮、半圆形外观,其交配能力较opaque状态的白念珠菌弱;而opaque细胞在显微镜下是伸长的圆柱状,胞内通常含有大液泡,体积也较white细胞大约3倍,而且细胞表面有较小的“pimples”(通过扫描电子显微镜检测到的未知生物作用的突起)(Anderson et al. 1990),在固体培养基上形成较为粗糙的灰色扁平状菌落,其交配效率大约是white细胞的100万倍.White和opaque细胞的细胞壁通透性也不同,在添加红色染料荧光桃红B(phloxine B)的固体平板上,white细胞形成白色菌落,而opaque细胞则吸收染料形成红色或粉红色菌落(Anderson & Soll 1987).菌丝的生长需要环境因子如37℃和血清等的持续刺激才能维持并且不能随着传代而遗传(Whiteway & Oberholzer 2004),而white和opaque两种形态细胞都可以稳定遗传并维持其原有形态几十代(Slutsky et al. 1987). ...
... 2.1.1 环境因子调控白念珠菌菌丝发生:人体的生理温度37℃是白念珠菌菌丝生长的最适温度(Whiteway & Oberholzer 2004).血清和GlcNAc是白念珠菌菌丝生长的两种诱导因子.血清诱导菌丝生长的成分可能是肽聚糖,人体自身不能合成肽聚糖,血清中的这种成分可能主要来源于肠道共生细菌代谢(Xu et al. 2008),而且GlcNAc是消化道黏膜和细菌细胞壁的组成成分(Levitt & Bond 1970).此外,高浓度CO2能够快速地诱导白念珠菌酵母形态向菌丝形态转换(Klengel et al. 2005),pH值在6.0-7.0时也有助于形成菌丝.细胞间的群体感应效应也会影响白念珠菌细胞的形态,高细胞密度时白念珠菌分泌的群体感应分子法尼醇(farnesol)对菌丝生长起明显的抑制作用,使得白念珠菌倾向于酵母形态的生长,而低细胞密度时更倾向于菌丝生长(Kumamoto 2005). ...
White-opaque switching in natural MTLa/alpha isolates of Candida albicans: evolutionary implications for roles in host adaptation, pathogenesis, and sex.
2
2013
... 在小鼠系统感染模型中,white细胞毒性最强,其次是opaque细胞,而gray细胞的毒性最弱.哺乳动物感染模型的初步研究表明opaque和gray细胞可能有对宿主上皮表面更高的适应能力(Xie et al. 2013;Tao et al. 2014),这说明白念珠菌面对不同的宿主环境,3种不同形态细胞表现不同的毒性特征. ...
... 2.2.2 转录水平调控白念珠菌white-gray-opaque转换:White-opaque转换与否的关键在于调控因子Wor1(white-opaque regulator 1)表达水平的高低(Huang et al. 2006;Srikantha et al. 2009).白念珠菌第5号染色体上存在与酿酒酵母MAT(mating-type)基因座类似的MTL(mating-type like)基因座,包含了MTLa1、MTLa2、MTLα1及MTLα2这4个基因(Magee & Magee 2000).MTL交配基因座不仅控制白念珠菌的交配,同时还控制white-opaque形态转换.自然界中绝大部分的白念珠菌是MTLa/α杂合型菌株,能合成Mtla1和Mtlα2形成Mtla1/α2异源复合物结合到WOR1的启动子区域抑制WOR1表达和opaque细胞形态的形成,最终抑制白念珠菌的交配(Zordan et al. 2006).因此自然界中很少有能进行交配的白念珠菌,长期以来国际上也一直认为白念珠菌只能以二倍体状态存在,不进行减数分裂和交配(Dolan & Fields 1991).随后才发现白念珠菌的交配效率与white-opaque形态转换密切相关,只有opaque形态细胞才能够进行高效率的交配(Park et al. 2013),而且这种交配也只能在MTL纯合的opaque细胞之间发生(Xie et al. 2013).不过后来发现,白念珠菌杂合型MTLa/α的OFR1突变株的opaque形态也可以分别与MTLα/∆型和MTLa/∆型opaque细胞进行交配(Sun et al. 2016). ...
Bacterial peptidoglycan triggers Candida albicans hyphal growth by directly activating the adenylyl cyclase Cyr1p.
2
2008
... 多种环境因子(如温度、血清、GlcNAc、低氧、CO2、pH、肽聚糖和氨基酸)都可通过信号转导通路组成的网络来调控白念珠菌的形态发生(Buffo et al. 1984;Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005;Xu et al. 2008). ...
... 2.1.1 环境因子调控白念珠菌菌丝发生:人体的生理温度37℃是白念珠菌菌丝生长的最适温度(Whiteway & Oberholzer 2004).血清和GlcNAc是白念珠菌菌丝生长的两种诱导因子.血清诱导菌丝生长的成分可能是肽聚糖,人体自身不能合成肽聚糖,血清中的这种成分可能主要来源于肠道共生细菌代谢(Xu et al. 2008),而且GlcNAc是消化道黏膜和细菌细胞壁的组成成分(Levitt & Bond 1970).此外,高浓度CO2能够快速地诱导白念珠菌酵母形态向菌丝形态转换(Klengel et al. 2005),pH值在6.0-7.0时也有助于形成菌丝.细胞间的群体感应效应也会影响白念珠菌细胞的形态,高细胞密度时白念珠菌分泌的群体感应分子法尼醇(farnesol)对菌丝生长起明显的抑制作用,使得白念珠菌倾向于酵母形态的生长,而低细胞密度时更倾向于菌丝生长(Kumamoto 2005). ...
Hgc1, a novel hypha-specific G1 cyclin-related protein regulates Candida albicans hyphal morphogenesis.
1
2004
... 2.2.1 转录水平调控白念珠菌菌丝形成:cAMP依赖的蛋白激酶A(cAMP-dependent protein kinase A,cAMP-PKA)是诱导菌丝形成的主要通路(Sudbery 2011).cAMP-PKA信号通路是真核生物中保守的信号传导途径,在酿酒酵母和白念珠菌的形态转换过程中起关键的调控作用(Hogan & Sundstrom 2009),能激活促进菌丝特异性基因表达的转录因子(Bockmuhl et al. 2001).CO2、GlcNAc和血清等环境因子诱导的白念珠菌菌丝生长主要通过这条途径发挥作用(Feng et al. 1999;Rocha et al. 2001;Klengel et al. 2005).Ras1是保守的GTP酶(small GTPase)蛋白,位于cAMP/PKA通路的上游,结合GTP后具有活性,过表达活性形式的RAS1(RAS1 G13V)能够促进菌丝生长(Takai et al. 2001).CYR1(CDC35)是白念珠菌中唯一腺苷酸环化酶编码基因(Rocha et al. 2001).胞外信号分子通过Ras1激活Cyr1,催化ATP转化为cAMP,导致胞内cAMP水平升高.cAMP结合PKA复合体的调节亚基Bcy1,使其与催化亚基解离,从而激活PKA的催化亚基,磷酸化并激活下游相应的转录因子,调控白念珠菌的菌丝生长(Bockmuhl & Ernst 2001;Cassola et al. 2004).该信号通路最终激活转录因子Efg1和Flo8等,调控菌丝特异性基因(HWP1、ECE1等)的表达,进而调控菌丝生长(Zheng et al. 2004). ...
Epigenetic properties of white-opaque switching in Candida albicans are based on a self-sustaining transcriptional feedback loop.
1
2006
... 2.2.2 转录水平调控白念珠菌white-gray-opaque转换:White-opaque转换与否的关键在于调控因子Wor1(white-opaque regulator 1)表达水平的高低(Huang et al. 2006;Srikantha et al. 2009).白念珠菌第5号染色体上存在与酿酒酵母MAT(mating-type)基因座类似的MTL(mating-type like)基因座,包含了MTLa1、MTLa2、MTLα1及MTLα2这4个基因(Magee & Magee 2000).MTL交配基因座不仅控制白念珠菌的交配,同时还控制white-opaque形态转换.自然界中绝大部分的白念珠菌是MTLa/α杂合型菌株,能合成Mtla1和Mtlα2形成Mtla1/α2异源复合物结合到WOR1的启动子区域抑制WOR1表达和opaque细胞形态的形成,最终抑制白念珠菌的交配(Zordan et al. 2006).因此自然界中很少有能进行交配的白念珠菌,长期以来国际上也一直认为白念珠菌只能以二倍体状态存在,不进行减数分裂和交配(Dolan & Fields 1991).随后才发现白念珠菌的交配效率与white-opaque形态转换密切相关,只有opaque形态细胞才能够进行高效率的交配(Park et al. 2013),而且这种交配也只能在MTL纯合的opaque细胞之间发生(Xie et al. 2013).不过后来发现,白念珠菌杂合型MTLa/α的OFR1突变株的opaque形态也可以分别与MTLα/∆型和MTLa/∆型opaque细胞进行交配(Sun et al. 2016). ...
Interlocking transcriptional feedback loops control white-opaque switching in Candida albicans.
1
2007
... 转录因子Czf1和Wor2可正向调节white-opaque转换,而转录因子Efg1可抑制white-opaque转换,维持细胞处于white状态.Czf1、Wor2和Efg1 3个转录因子与Wor1共同组成一个反馈环调控白念珠菌white-opaque形态转换,Wor1处于这个调控网络的中心地位.Wor1不仅能够结合到自身启动子区域调控自身表达,还能够结合到EFG1、CZF1和WOR2的启动子区域,抑制EFG1表达,激活CZF1和WOR2表达.因此,当WOR1表达增加时会抑制EFG1的表达并促进opaque的状态,而WOR1表达下降时会引起对EFG1表达的去抑制作用(Srikantha et al. 2000;Vinces & Kumamoto 2007;Zordan et al. 2007;Ramirez-Zavala et al. 2008).此外染色体修饰也可以影响white-opaque转换的效率,例如降低组蛋白去乙酰化水平能促进细胞从white状态转换为opaque状态(Morschhauser 2010). ...






{kind=link}
{kind=link}
{kind=link}
{kind=link}