


小菇属Mycena (Pers.) Roussel是小菇科最大的属,全世界至少有500种,绝大多数为腐生菌(Kirk et al. 2008)。近150年来,随着美洲、欧洲、大洋洲、亚洲的专著先后出版,各大洲描述的小菇属物种达200余种(Smith 1947;Robich 2003;Kirk et al. 2008;Aravindakshan 2015;Aronsen & Læssøe 2016)。小菇属真菌是白色孢子伞菌的重要组成部分,具有重要的生态、经济和科研价值(魏玉莲和戴玉成2004;戴玉成和杨祝良2008;戴玉成等2010),近年来,该属真菌在兰科植物资源保育等领域提供了新理论依据(Guo et al. 1997;Boa 2004;Peters & Spiteller 2007;高越等 2019)。20世纪,小菇属以Maas Geesteranus(1992a,1992b)为代表的分类系统为主,随着蘑菇目大系统框架的构建,成立小菇科的观点被认可和采纳(Moncalvo et al. 2002;Matheny 2006;娜琴 2019)。由于小菇属物种数量多、个体小、种间差异不明显,加之认知水平不完善、取样量严重不足等诸多原因,导致小菇属物种识别不清,属下系统学框架极不清晰。
过去50年,小菇属在国内共记录了76个名称,发现6个新种,16个中国新记录种(戴芳澜 1979;毕志树等 1984;Bi & Li 1986;何显1994;Guo et al. 1999;李玉等 2015)。从整体上看,国内研究人员对小菇属开展过一些研究工作,但以经典分类为主,缺乏对系统演化规律的探讨。作者在小菇属的形态分类和分子系统学研究中用多基因建树等方法进行了尝试(Na & Bau 2018,2019a ,2019b;娜琴 2019)。总体上看,国内对小菇属真菌物种多样性及演化历史的认知还有较大局限。
本研究的小菇属10个中国新记录种隶属于担孢子淀粉质和非淀粉质的7个组。Redhead(2012)将非淀粉质担孢子的香菌组划分在Atheniella中,由于其观点缺乏系统学证据而并未发表。另外,前期研究结果支持了棘刺组作为小菇属属下等级的结果,Ampariona heteracantha作为Mycena heteracantha的异名(娜琴 2019;Na & Bau 2019b)。
1 材料与方法
1.1 材料
研究材料为2016至2018年间在内蒙古、辽宁、吉林、黑龙江、湖南、浙江、安徽、福建、四川、云南、西藏等11个省、自治区采集获得。硅胶干燥法制作研究标本及分子材料。
1.2 方法
采用传统的形态学分类方法,显微特征观察以5% KOH作为浮载剂,必要时用1%刚果红染色,随机测量40个成熟担孢子,测量数值小数点后保留一位。担子、褶缘囊状体、侧生囊状体(若有)、柄生囊状体等特征均随机选取20个测量并记录形状、大小、有无内含物。
试剂盒法对所制作分子材料提取DNA,采用25μL PCR扩增体系和Touchdown程序,引物选用ITS1和ITS4(White et al. 1990)。Mrmodeltest 2.3选择最优模型(Posada & Crandall 1998)。分别构建贝叶斯和最大似然系统发育树(Stamatakis et al. 2004;Ronquist et al. 2012)。
2 结果与分析
2.1 系统学
表1 系统发育分析中自测和GenBank下载的ITS序列信息
Table 1
| 序号 Sample | 种名 Species | 凭证号 Voucher | GenBank号 GenBank accession No. | 来源地 Locality |
|---|---|---|---|---|
| 1 | Mycena adonis (Bull.) Gray | HMJAU 43008 | MK309769 | China |
| 2 | Mycena adscendens Maas Geest. [=Mycena tenerrima (Berk.) Quél.] | Aronsen120803 | KT900140 | Sweden |
| 3 | Mycena adscendens Maas Geest. | Orstadius329-05 | KT900141 | Sweden |
| 4 | Mycena adscendens Maas Geest. | Aronsen061119 | KT900142 | Sweden |
| 5 | Mycena adscendens Maas Geest. | Aronsen120826/1 | KT900143 | Sweden |
| 6 | Mycena amabillissima (Peck) Sacc. | AFTOL-ID 1686 | DQ490644 | USA |
| 7 | Mycena amicta (Fr.) Quél. | 4745-HRL | KJ705188 | Canada |
| 8 | Mycena amicta (Fr.) Quél. | 189f | JF908394 | Italy |
| 9 | Mycena amicta (Fr.) Quél. | CBS 257.53 | MH857184 | Nertherlands |
| 10 | Mycena amygdalina (Pers.) Singer | HMJAU 43629 | MT497544 | China |
| 11 | Mycena amygdalina (Pers.) Singer | HMJAU 43700 | MT497545 | China |
| 12 | Mycena arcangeliana Bres. | 252b | JF908401 | Italy |
| 13 | Mycena arcangeliana Bres. | 252f | JF908402 | Italy |
| 14 | Mycena aurantiidisca (Murrill) Murrill | DQ384585 | DQ384585 | Unpublished |
| 15 | Mycena aurantiidisca (Murrill) Murrill | HMJAU 43811 | MT497546 | China |
| 16 | Mycena chlorophos (Berk. & M.A. Curtis) Sacc. | ACL093 | KJ206970 | Malaysia |
| 17 | Mycena chlorophos (Berk. & M.A. Curtis) Sacc. | ACL257 | KJ206983 | Malaysia |
| 18 | Mycena chlorophos (Berk. & M.A. Curtis) Sacc. | ACL271 | KJ206986 | Malaysia |
| 19 | Mycena cinerella (P. Karst.) P. Karst. | 173 | MF926553 | Russia |
| 20 | Mycena cinerella (P. Karst.) P. Karst. | Aronsen051014 | KT900146 | Sweden |
| 21 | Mycena citrinomarginata Gillet | AD4TN | KU973883 | Tunisia |
| 22 | Mycena clavata (Peck) Redhead | PRM:935333 | LT671448 | Czech |
| 23 | Mycena clavata (Peck) Redhead | PRM:m935283 | LT671449 | Czech |
| 24 | Mycena clavicularis (Fr.) Gillet | 615i | JF908466 | Italy |
| 25 | Mycena filopes (Bull.) P. Kumm. | 3782 | KJ705175 | Unpublished |
| 26 | Mycena filopes (Bull.) P. Kumm. | 287f | JF908410 | Italy |
| 27 | Mycena flavoalba (Fr.) Quél. | HMJAU 43764 | MT497547 | China |
| 28 | Mycena floridula (Fr.) Quél. | HMJAU 43193 | MK309770 | China |
| 29 | Mycena floridula (Fr.) Quél. | HMJAU 43213 | MK309771 | China |
| 30 | Mycena floridula (Fr.) Quél. | HMJAU 43613 | MK309772 | China |
| 31 | Mycena heteracantha (Singer) Desjardin | HMJAU 43709 | MK309785 | China |
| 32 | Mycena heteracantha (Singer) Desjardin | HMJAU 43711 | MK309786 | China |
| 33 | Mycena heteracantha (Singer) Desjardin | HMJAU 43716 | MK309787 | China |
| 34 | Mycena hiemalis (Osbeck) Quél. | HMJAU 43620 | MK309790 | China |
| 35 | Mycena hiemalis (Osbeck) Quél. | 45d | JF908444 | Italy |
| 36 | Mycena illuminans Henn. | ACCL175 | KJ206976 | Malaysia |
| 37 | Mycena illuminans Henn. | ACL212 | KJ206980 | Malaysia |
| 38 | Mycena leaiana (Berk.) Sacc. | 1028 | JF908376 | Italy |
| 39 | Mycena leaiana (Berk.) Sacc. | CNH03(TENN) | MF686520 | Unpublished |
| 40 | Mycena leaiana (Berk.) Sacc. | NAWA2017-132 | MH979290 | Unpublished |
| 41 | Mycena luteopallens (Peck) Peck | 430287 | AB512313 | Unpublished |
| 42 | Mycena maculata P. Karst. | HMJAU 43009 | MK309791 | China |
| 43 | Mycena maculata P. Karst. | HMJAU 43111 | MK309792 | China |
| 44 | Mycena maurella Robich | 314a | JF908413 | Italy |
| 45 | Mycena maurella Robich | 314h | JF908414 | Italy |
| 46 | Mycena meliigena (Berk. & Cooke) Sacc. | 39 | JF908423 | Italy |
| 47 | Mycena meliigena (Berk. & Cooke) Sacc. | 39d | JF908429 | Italy |
| 48 | Mycena metata (Secr. ex Fr.) P. Kumm. | 313b | JF908412 | Italy |
| 49 | Mycena murina (Murrill) Murrill | F14062 | AF335444 | Unpublished |
| 50 | Mycena murina (Murrill) Murrill | OT10 | KJ093469 | Unpublished |
| 51 | Mycena olida Bres. | 292v | JF908411 | Italy |
| 52 | Mycena olida Bres. | WU 32745 | KF499358 | Unpublished |
| 53 | Mycena olida Bres. | KA13-1261 | KR673718 | Korea |
| 54 | Mycena olida Bres. | BIOUG24046-C02 | KT695358 | Unpublished |
| 55 | Mycena pearsoniana Dennis ex Singer | TENN61544 | JN182199 | Italy |
| 56 | Mycena pearsoniana Dennis ex Singer | TENN61384 | JN182200 | Italy |
| 57 | Mycena pelianthina (Fr.) Quél. | 108b | JF908379 | Italy |
| 58 | Mycena pelianthina (Fr.) Quél. | 108f | JF908380 | Italy |
| 59 | Mycena plumbea P. Karst. | JN198391 | JN198391 | Unpublished |
| 60 | Mycena polygramma (Bull.) Gray | 439b | JF908433 | Italy |
| 61 | Mycena polygramma (Bull.) Gray | 439f | JF908434 | Italy |
| 62 | Mycena pura (Pers.) P. Kumm. | TENN65043 | JN182202 | USA |
| 63 | Mycena purpureofusca (Peck) Sacc. | F19748 | HQ604766 | Canada |
| 64 | Mycena rosella (Fr.) P. Kumm. | 938a | JF908488 | Italy |
| 65 | Mycena rosella (Fr.) P. Kumm. | Champ-21 | KX449424 | Spain |
| 66 | Mycena rubromarginata (Fr.) P. Kumm. | 407q | JF908430 | Italy |
| 67 | Mycena rubromarginata (Fr.) P. Kumm. | TL-12780 | KX513845 | USA |
| 68 | Mycena sanguinolenta (Alb. & Schwein.) P. Kumm. | 136f | JF908390 | Italy |
| 69 | Mycena seminau A.L.C. Chew & Desjardin | ACL136 | KF537250 | Malaysia |
| 70 | Mycena seminau A.L.C. Chew & Desjardin | ACL308 | KF537252 | Malaysia |
| 71 | Mycena semivestipes (Peck) A.H. Sm. | TENN61770 | FJ596888 | Unpublished |
| 72 | Mycena seynii Quél. | 71l | JF908469 | Italy |
| 73 | Mycena seynii Quél. | 71h | JF908470 | Italy |
| 74 | Mycena silvae-nigrae Maas Geest. & Schwöbel | 515 | JF908452 | Italy |
| 75 | Mycena silvae-nigrae Maas Geest. & Schwöbel | CC13-12 | KF359604 | USA |
| 76 | Mycena speirea (Fr.) Gillet | HMJAU 43520 | MT497548 | China |
| 77 | Mycena speirea (Fr.) Gillet | HMJAU 43694 | MT497549 | China |
| 78 | Mycena speirea (Fr.) Gillet | 218n | JF908396 | Italy |
| 79 | Mycena speirea (Fr.) Gillet | PRM:922296 | LT671445 | Czech |
| 80 | Mycena strobilinoidea Peck | 151e | JF908393 | Italy |
| 81 | Mycena supina (Fr.) P. Kumm. | 128a | JF908388 | Italy |
| 82 | Mycena tenax A.H. Sm. | p187i | EU669224 | USA |
| 83 | Mycena tenax A.H. Sm. | OSC 113746 | EU846251 | USA |
| 84 | Mycena viridimarginata P. Karst. | 104h | JF908378 | Italy |
| 85 | Mycena vulgaris (Pers.) P. Kumm. | 447h | JF908435 | Italy |
| 86 | Mycena vulgaris (Pers.) P. Kumm. | 3781 | KJ705177 | Canada |
| 87 | Mycena zephirus (Fr.) P. Kumm. | HMJAU 43106 | MT497550 | China |
| 88 | Mycena zephirus (Fr.) P. Kumm. | KA13-1265 | KR673722 | Korea |
| 89 | Xeromphalina campanella (Batsch) Kühner & Maire | TFB7283A | KM024575 | Sweden |
| 90 | Xeromphalina campanella (Batsch) Kühner & Maire | TFB14487 | KP835678 | Czech |
Note: The newly generated sequences in this study were marked in bold.
注:本研究涉及的自测序列以粗体表示
图1
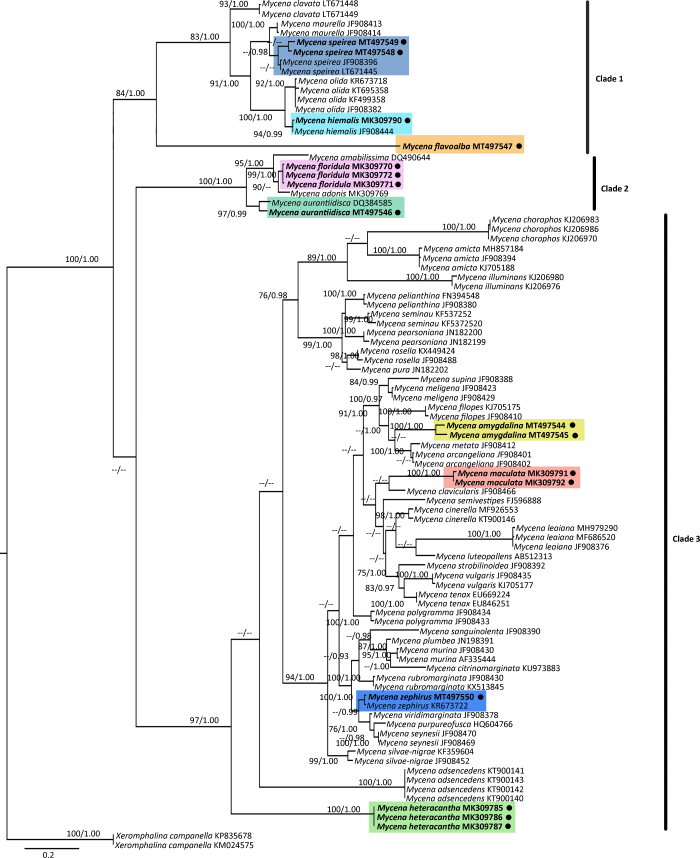
图1
基于ITS序列构建的小菇属系统发育树(ML≥75;BPP≥0.95)
Fig. 1
Phylogenetic tree of Mycena based on ITS (ML≥75; BPP≥0.95).
2.2 形态学
Mycena amygdalina (Pers.) Singer, Persoonia 2(1): 6 (1961).
≡Agaricus amygdalinus Pers., Mycol. eur. (Erlanga) 3: 255 (1828).
菌盖直径5-10mm,斗笠形,中央具乳突,灰白色、灰色至深灰色,中央深灰色,表面被白粉霜,干,具条纹,形成浅沟槽。菌肉灰白色,易碎,薄。味道不明显,气味强烈碱味。菌褶白色、灰白色,直生,垂齿状。菌柄44-62×0.5-1.0mm,中空,圆柱形,脆骨质,上部灰白色,中部灰色,下部灰紫色,初期表面具粉霜,老后渐光滑,基部具白色绒毛。
图2

图2
新记录种野外生境照片
A,B:碱味小菇;C,D:橙盖小菇;E,F:黄白小菇;G,H:粉黄小菇;I:绒柄小菇;J-L:异刺小菇;M,N:冬生小菇;O:红斑小菇;P,Q:绣线菊小菇;R,S:粉被小菇. 标尺:A-S=10mm
Fig. 2
Photographs of new record sepcies in the wild.
A, B: Mycena amygdalina; C, D: M. aurantiidisca; E, F: M. flavoalba; G, H: M. floridula; I: M. flos-nivium; J-L: M. heteracantha; M, N: M. hiemalis; O: M. maculata; P, Q: M. speirea; R, S: M. zephirus. Bars: A-S=10mm.
担孢子7.1-8.6×2.7-4.0μm,Q=1.9-2.6,圆柱形至长圆柱形,光滑,无色,淀粉质,薄壁。担子22-30×6-8μm,棒状,具4个小梗。褶缘囊状体20-37×12-17μm,圆柱形、棒状、倒卵圆形、倒梨形,上部具密集圆柱状瘤疣突,无色。褶侧囊状体19-31×11-16μm,形状与褶缘囊状体相似。菌盖表皮表面密被棒状瘤疣突起,1.3-4.7×1.0-1.6μm。菌柄皮层表面形成浓密圆柱状瘤疣状突起,1.0-4.3×0.7-1.5μm。具锁状联合。
生境:夏秋季散生于落叶松、红松等针叶林枯枝落叶层。
中国分布:内蒙古、吉林、黑龙江、台湾。
研究标本:内蒙古:根河市汗马国家级自然保护区,娜琴,2016年9月3日,HMJAU 43349;鄂温克旗辉河国家级自然保护区,娜琴,2016年9月4日,HMJAU 43354;呼伦贝尔市满归镇伊克萨玛国家森林公园,娜琴,2017年7月30日,HMJAU 43629,HMJAU 43630,黄梅,2018年7月24日,HMJAU 43400;呼伦贝尔市得耳布尔镇康达岭林场,娜琴,2017年8月21日,HMJAU 43700。吉林:延边朝鲜族自治州安图县二道白河镇科学院后山,2016年6月15日,HMJAU 43024,HMJAU 43043;延边朝鲜族自治州安图县二道白河镇老山门,娜琴,2016年8月27日,HMJAU 43314。黑龙江:漠河县松苑,娜琴,2017年8月25日,HMJAU 43392;塔河县十八站鄂祖乡,娜琴,2017年8月28日,HMJAU 43393。
图3

图3
碱味小菇
A:担子体;B:担孢子;C:担子;D:褶侧囊状体;E:褶缘囊状体;F:菌盖表皮;G:菌柄皮层. 标尺:A=10mm,B-G=10μm
Fig. 3
Mycena amygdalina.
A: Basidiomata; B: Basidiospores; C: Basidia; D: Pleurocystidia; E: Cheilocystidia; F: Pileipellis; G: Stipitipellis. Bars: A=10mm, B-G=10μm.
讨论:Kühner(1938)和Smith(1947)因该种具有明显味道而命名为M. iodiolens Lundell.。Singer(1961)提出了新组合M. amygdalina (Pers.) Sing. comb nov.,M. iodiolens处理为异名。
M. amygdalina在我国最早由Chou(2004)等作为台湾新记录种发表,但欧洲学者认为M. amygdalina应合并在M. filopes complex中,并不赞同其为独立的物种。欧洲记载的M. filopes担孢子9-11.5×5.5-6.5μm,Qav=1.7,椭圆形,而自采材料为圆柱形,形状差异显著。
M. vitilis (Fr.) Quél.与M. amygdalina形态学特征相似,但前者无明显气味,担孢子长椭圆形,褶缘囊状体烧瓶形,顶端具2-3个分支,无褶侧囊状体。另外,M. arcangeliana Bres在野外与本种易混淆,但M. arcangeliana强烈碘酒味,菌盖橄榄绿色,褶缘囊状体及褶侧囊状体较大,担孢子为椭圆形至长椭圆形(Aronsen & Læssøe 2016)。
Mycena aurantiidisca (Murrill) Murrill, Mycologia 8(4): 220 (1916).
≡Atheniella aurantiidisca (Murrill) Redhead, Moncalvo, Vilgalys, Desjardin & B.A. Perry, Index Fungorum 14: 1 (2012).
菌盖直径2-13mm,幼时凸镜形、半球形,成熟后圆锥形,中央突起,橙黄色、橙色至橙红色,成熟后褪色至淡橙黄色、黄白色,边缘颜色较浅,淡黄色,表面透明状条纹,边缘微波浪状。菌肉白色,薄。气味和味道不明显。菌褶白色、黄白色,直生至弯生,垂齿状,具1-2小褶片。菌柄21-92×1.0-2.0mm,中空,圆柱形,脆骨质,白色、黄白色,上部表面被粉霜,成熟后近光滑,基部少量白色菌丝体。
担孢子6.0-7.3×2.9-3.7μm,Q=2.0-2.7,长椭圆形、圆柱形,光滑,无色,非淀粉质。担子21-19×6-8μm,棍棒状,具4个小梗。褶缘囊状体23-40×7-11μm,拟纺锤形、烧瓶形,光滑。褶侧囊状体稍小,20-36×6-10μm。菌盖表皮菌丝表面被圆柱形或棒状瘤疣突,无色。柄生囊状体12-20×6-10μm,棒状、纺锤形、近球形至头部膨大,光滑。具锁状联合。
生境:夏秋季散生、群生于云杉、华山松等针叶林腐木苔藓层。
中国分布:四川、云南、西藏。
研究标本:云南:迪庆藏族自治州香格里拉普达措国家公园,娜琴,2018年8月15日,HMJAU 43808。西藏:林芝市波密县多东寺,娜琴,2018年8月18日,HMJAU 43811。
讨论:Mycena aurantiidisca最初被认知为M. acicula的一个变型,二者间的宏观形态特征高度相似,但M. acicula菌盖橙红色,成熟后并不发生褪色,菌盖表皮由泡状细胞组成,褶缘囊状体和褶侧囊状体头部钝圆,另外,M. acicula仅生于在阔叶林的枯枝落叶层,而M. aurantiidisca只发现在针叶林中有分布(Murrill 1916;Maas Geesteranus 1990)。M. oregonensis菌盖颜色与M. aurantiidisca不易分辨,但前者菌褶和菌柄橙黄色,褶缘囊状体及柄囊体具黄色内含物。M. leptophylla区别于M. aurantiidisca的特征为褐色菌柄、近球形担孢子、棒状柄囊体。
图4
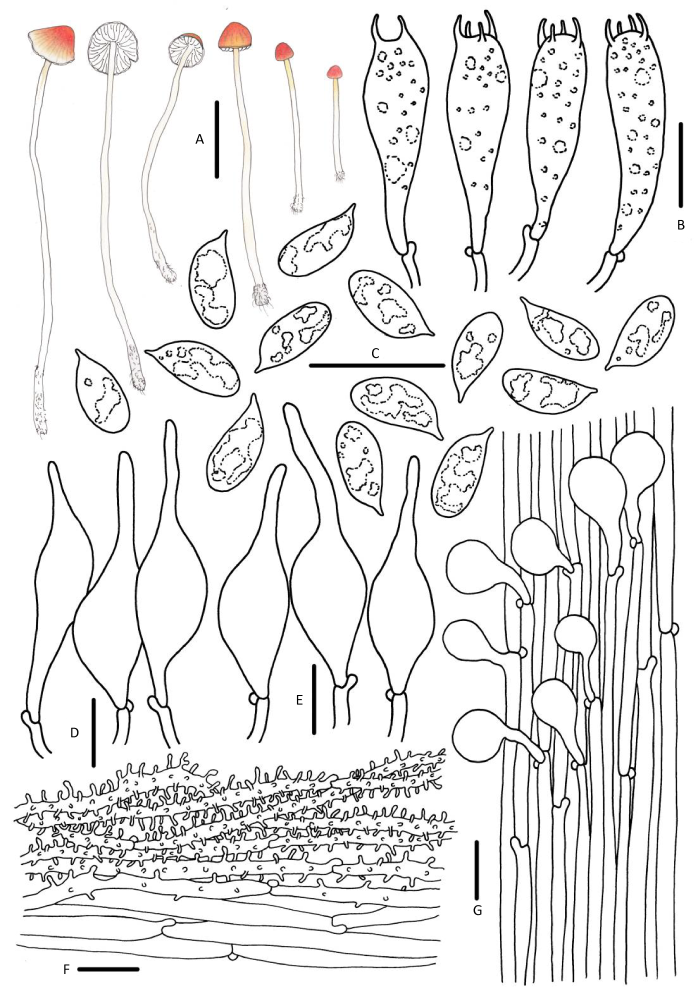
图4
橙盖小菇
A:担子体;B:担子;C:担孢子;D:褶缘囊状体;E:褶侧囊状体;F:菌盖表皮;G:菌柄皮层. 标尺:A=10mm,B-G=10μm
Fig. 4
Mycena aurantiidisca.
A: Basidiomata; B: Basidia; C: Basidiospores; D: Cheilocystidia; E: Pleurocystidia; F: Pileipellis; G: Stipitipellis. Bars: A=10mm, B-G=10μm.
Mycena flavoalba (Fr.) Quél., Mém. Soc. Émul. Montbéliard, Sér. 2 5: 103 (1872).
≡Agaricus flavoalbus Fr. 1838.
菌盖直径4-21mm,半球形、钟形,中央钝圆突起,成熟后渐平展,有时中央下凹,黄白色、淡黄色,边缘白色,光滑,具透明状条纹。菌肉白色,易碎。无明显气味,味道温和。菌褶白色,弯生,垂齿状。菌柄25-62×1.0-2.5mm,中空,圆柱形,脆骨质,白色,上部微被粉霜,向下渐光滑,基部具少量白色绒毛。
图5
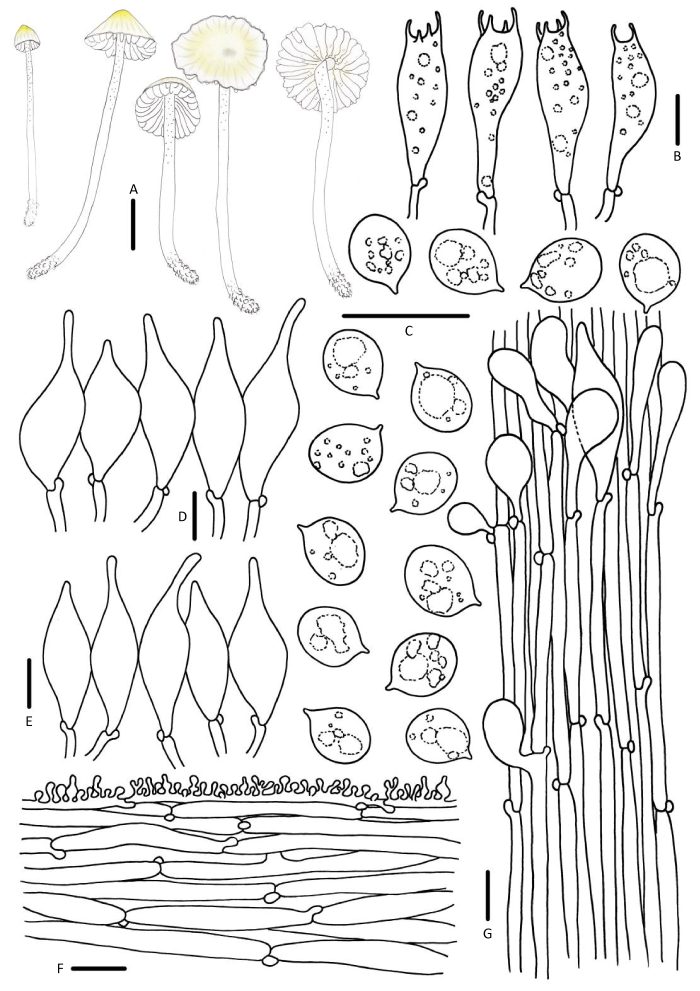
图5
黄白小菇
A:担子体;B:担子;C:担孢子;D:褶缘囊状体;E:褶侧囊状体;F:菌盖表皮;G:菌柄皮层. 标尺:A=10mm,B-G=10μm
Fig. 5
Mycena flavoalba.
A: Basidiomata; B: Basidia; C: Basidiospores; D: Cheilocystidia; E: Pleurocystidia; F: Pileipellis; G: Stipitipellis. Bars: A=10mm, B-G=10μm.
担孢子5.8-7.5×4.5-6.9μm,Q=1.1-1.3,宽椭圆形、近球形,无色,光滑,非淀粉质。担子22-30×5-7μm,棒状,具2或4个小梗。缘生囊状体25-56×6-14μm,细颈烧瓶形,光滑,无色。侧生囊状体较小,21-49×5-13μm。菌盖表皮菌丝表面棒状瘤疣突起,3.5-5.6×1.8-2.9μm。柄生囊状体棍棒状、纺锤形、烧瓶形、近球形,12-47× 10-15μm。具锁状联合。
生境:夏秋季单生、散生于草坪上。
中国分布:浙江、安徽、福建、四川。
研究标本:浙江:杭州市浙江大学紫金港校区校园,娜琴,2018年7月1日,HMJAU 43755。安徽:黄山市国元大酒店门前,娜琴,2018年7月3日,HMJAU 43764。福建:福州市福州大学校园内,曾辉,2018年8月26日,HMJAU 43848。四川:甘孜藏族自治州康定市木格措风景区,娜琴,2017年8月21日,HMJAU 43676,HMJAU 43682。
讨论:有学者认为Mycena flavoalba因其非淀粉质孢子这一特征应划分在Hemimycena中,但并未公开发表,Maas Geesteranus(1990)提出的小菇属的主流分类系统将M. flavoalba隶属于sect. Adonideae。Favre(1948)和Robich(2003)均发现了具苦味的新变种在欧洲有分布,即M. flavoalba var. amara。欧洲学者Aronsen & Læssøe(2016)并不赞同M. flavoalba种下等级的划分,而是作为广义M. flavoalba来认知。
中国亚热带地区的M. flavoalba材料担孢子为宽椭圆形至近球形,与欧洲温带地区描述的长椭圆形担孢子差异较大,但其他形态学特征均吻合。
前期研究的系统发育树结果揭示了M. flavoalba与sect. Adonideae存在一定的遗传距离,并不认同M. flavoalba隶属于sect. Adonideae。但中国研究材料采自亚热带,并未收集到北温带的材料,因此系统发育树结果并不完善,暂时将M. flavoalba保留在sect. Adonideae中。由于缺乏系统学结果的支持,M. flavoalba同上述M. aurantiidisca均保留在小菇属中,暂不采用Atheniella flavoalba作为该种学名(娜琴 2019)。
Mycena floridula (Fr.) Quél., Bull. Soc. bot. Fr. 23: 325 (1877) [1876].
≡Agaricus floridulus Fr., Epicr. syst. mycol. (Upsaliae): 94 (1838) [1836-1838].
菌盖直径3-16mm,初期半球形、凸镜形,老后渐平展,中央钝圆突起,淡粉色、粉色、粉黄色,中央及边缘黄白色、乳白色,表面光滑,湿时黏。菌肉白色。无明显气味与味道。菌褶白色或淡粉色,直生,垂齿状。菌柄25-72×1.5-3.0mm,中空,圆柱形,脆骨质,白色、黄白色,幼时被白色细小绒毛,老后近光滑,基部具少量白色绒毛。
图6
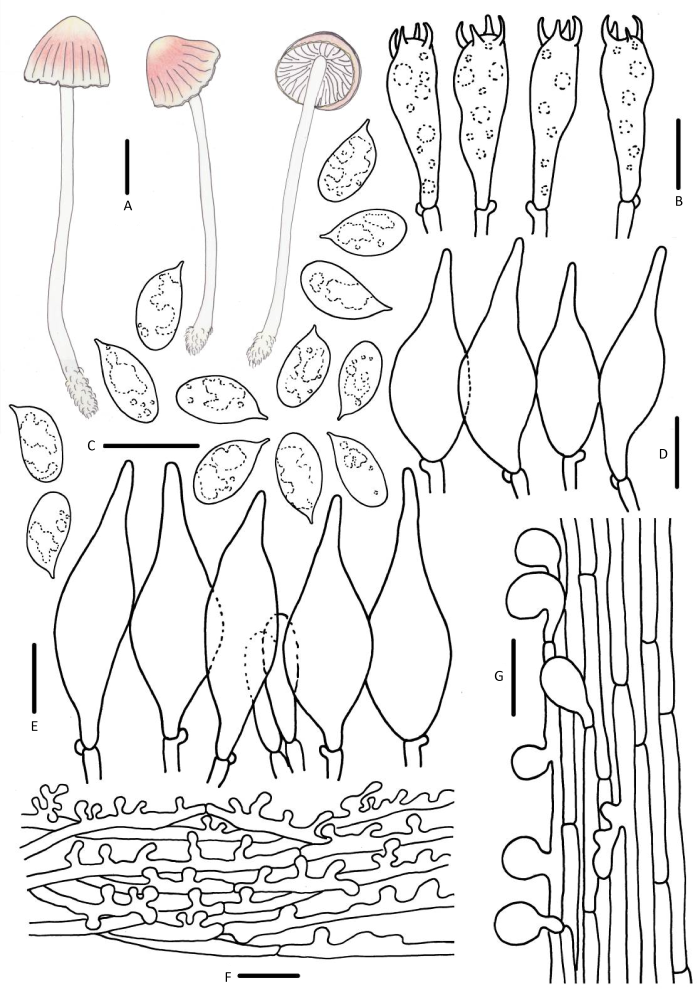
图6
粉黄小菇
A:担子体;B:担子;C:担孢子;D:褶侧囊状体;E:褶缘囊状体;F:菌盖表皮;G:菌柄皮层. 标尺:A=10mm,B-G=10μm
Fig. 6
Mycena floridula.
A: Basidiomata; B: Basidia; C: Basidiospores; D: Pleurocystidia; E: Cheilocystidia; F: Pileipellis; G: Stipitipellis. Bars: A=10mm, B-G=10μm.
担孢子5.8-8.2×3.5-4.5μm,Q=1.8-2.2,长椭圆形、圆柱形,无色,光滑,非淀粉质。担子20-30×5-8μm,棒状,具4个小梗。褶缘囊状体37-54×8-13μm,拟纺锤形、细颈烧瓶形,无色。褶侧囊状体较小,29-38×7-11μm。菌盖表皮表面不规则瘤疣突起,7-15×6-8μm。菌柄皮层菌丝具近球形,3-8×1-6μm,光滑。具锁状联合。
生境:夏秋季单生于落叶松等针叶林枯枝落叶层上。
中国分布:内蒙古、吉林、黑龙江。
研究标本:内蒙古:呼伦贝尔市满归镇伊克萨玛国家森林公园,娜琴,2016年8月12日,HMJAU 43193,2017年7月30日,HMJAU 43633;呼伦贝尔市满归镇孟贵线12km处,娜琴,2017年7月29日,HMJAU 43613。吉林:延边朝鲜族自治州安图县二道白河镇老山门,娜琴,2016年8月27日,HMJAU 43313。黑龙江:双河国家级自然保护区,娜琴,2016年8月15日,HMJAU 43213。
讨论:欧洲早期的文献将M. floridula认知为M. flavoalba的粉色变型,二者在显微结构并无差异(Kühner 1938)。Maas Geesteranus(1991)认为M. floridula 应为独立的物种,强调了M. floridula与M. flavoalba菌盖颜色的差异。Aronsen & Læssøe(2016)支持早期欧洲学者的观点,认为M. floridula应合并在M. flavoalba中。
相近种M. adonis(≡Atheniella adonis)菌盖和菌柄无明显黄色,老后菌盖仅褪为白色,担孢子椭圆形至长椭圆形可以很好地区分于M. floridula(Maas Geesteranus 1991)。基于3个基因片段构建的系统发育树显示M. floridula形成单独的分支,支持其为一个独立的物种(娜琴 2019)。
Mycena flos-nivium Kühner, Bull. Soc. nat. Oyonnax 6: 71 (1952).
菌盖直径22-45mm,圆锥形、斗笠形,老后钟形,中央钝圆突起,黄褐色、淡褐色、褐色,中央暗褐色,边缘米黄色,初期微被白色细小绒毛,后光滑,具透明状条纹。菌肉白色,薄。气味不明显,味道淡淀粉味。菌褶白色至灰白色,直生,垂齿状,窄,褶间具横脉。菌柄34-66× 1.5-4.0mm,中空,圆柱形,脆骨质,光滑,白色、污白色、灰白色,向下渐深至黄褐色,基部被密集白色菌丝体。
图7
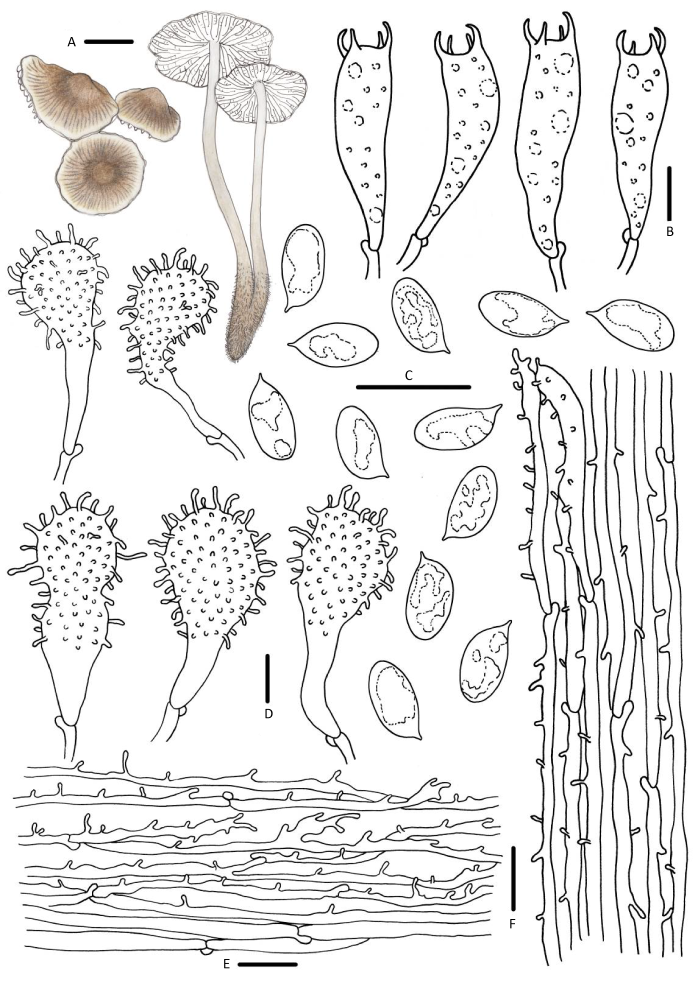
图7
绒柄小菇
A:担子体;B:担子;C:担孢子;D:褶缘囊状体;E:菌盖表皮;F:菌柄皮层. 标尺:A=10mm,B-F=10μm
Fig. 7
Mycena flos-nivium.
A: Basidiomata; B: Basidia; C: Basidiospores; D: Cheilocystidia; E: Pileipellis; F: Stipitipellis. Bars: A=10mm, B-F=10μm.
担孢子7.9-9.5×4.3-4.8μm,Q=1.9-2.2,长椭圆形、圆柱形,无色,光滑,淀粉质。担子23-31×6-8μm,棍棒状,具4个小梗。褶缘囊状体棒状或顶端稍膨大,21-65×7-12μm,具密集不规则瘤突。褶侧囊状未见。菌盖表皮菌丝偶见疣突或圆柱状突起,1.5-2.8×1.0-1.2μm。菌柄皮层偶见圆柱状疣突,1.8-7.5×0.6-1.1μm。具锁状联合。
生境:夏秋季单生、散生于红松、冷杉、云杉、红豆杉等针叶林腐木上。
中国分布:吉林。
研究标本:吉林:延边朝鲜族自治州安图县二道白河镇科学院后山,娜琴,2016年6月15日,HMJAU 43019,HMJAU 43026;和龙市甄峰岭,娜琴,2016年6月17日,HMJAU 43097。
讨论:Mycena flos-nivium广泛分布在欧洲大陆,Kühner(1938)最早命名该种,但仅记录了简单的宏观特征,Maas Geesteranus(1985)基于Kühner(1938)的凭证标本提供了详细的形态学描述。
Maas Geesteranus(1985)提出的分类系统中,M. plumipes (Kalchbr.) P.-A. Moreau和M. Silvae-nigrae Maas Geest. & Schwöbel与M. Flos-nivium相似,但M. plumipes菌柄褐色,担孢子宽椭圆形,褶缘囊状体两型,纺锤形且光滑、顶端具有指状分枝。M. Silvae-nigrae菌盖褐色,黏,担孢子长椭圆形,褶缘囊状体光滑,菌盖表皮具凝胶层(Robich 2003;Aronsen & Læssøe 2016)。
Mycena heteracantha (Singer) Desjardin, Biblthca Mycol. 159: 18 (1995).
菌盖直径1.8-4.2mm,斗笠形、半球形、钟形,幼时中央明显钝圆突起,白色,表面具密集白色麸状、皮屑状绒毛,边缘具稀疏“砂糖”状绒毛,半透明状条纹,形成浅沟槽。菌肉白色,薄。气味和味道不明显。菌褶白色,直生至弯生,垂齿状,边缘老后常开裂。菌柄11-35×0.5-0.8mm,中空,圆柱形,细,脆骨质,白色,表面密被白色绒毛,基部形成绒毛状圆盘。
图8
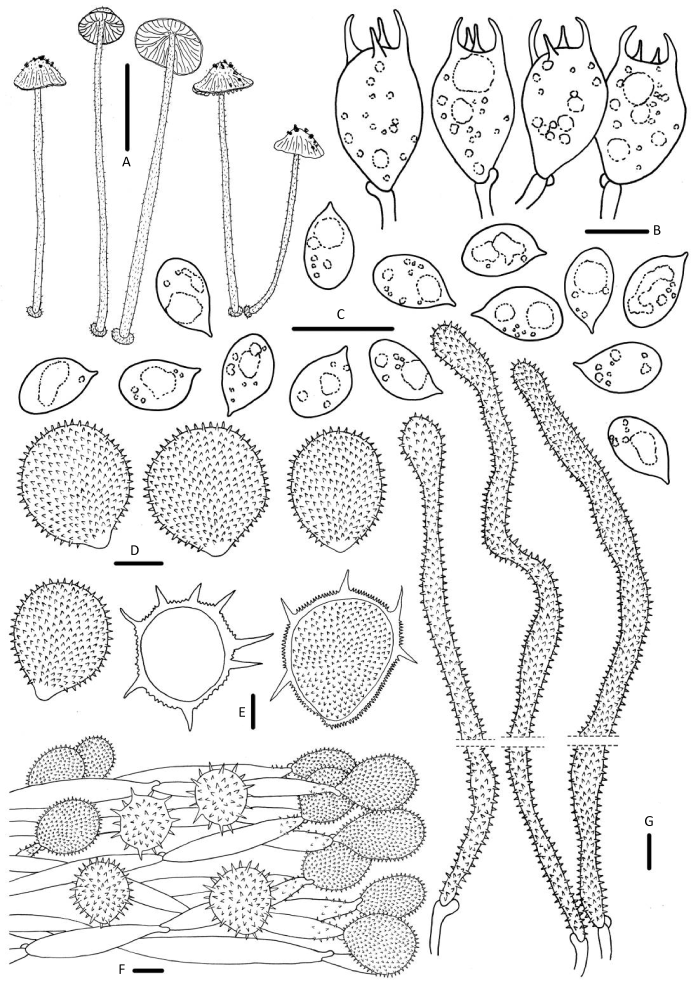
图8
异刺小菇
A:担子体;B:担子;C:担孢子;D:褶缘囊状体;E:菌盖表皮长刺状特化细胞;F:菌盖表皮;G:柄生囊状体. 标尺:A=10mm,B-G=10μm
Fig. 8
Mycena heteracantha.
A: Basidiomata; B: Basidia; C: Basidiospores; D: Cheilocystidia; E: Cherocytes; F: Pileipellis; G: Caulocystidia. Bars: A=10mm, B-G=10μm.
担孢子7.9-9.7×5.5-6.1μm,Q=1.4-1.8,椭圆形,光滑,薄壁,无色,淀粉质。担子15-21× 9-13μm,棍棒状,具4个小梗。褶缘囊状体泡囊状、倒卵圆形、倒梨形,23-34×19-27μm,具密集刺状疣突。褶侧囊状体缺失。菌盖表皮表面具被长刺状特化细胞cherocytes,23-56×21-45μm,近球形且基部花梗状,末端分化细胞acanthocysts倒梨形、泡囊形,18-63×16-49μm。柄囊体110-307×8-12μm,圆柱形、长棍棒状,表面密集刺状突起。具锁状联合。
生境:夏秋季散生、群生于樟树、栎等阔叶林腐木或枯枝落叶层上。
中国分布:浙江、湖南。
研究标本:浙江:浙江大学紫金港校区,娜琴,2018年7月1日,HMJAU 43756。湖南:长沙市岳麓山风景区,娜琴,2018年5月13日,HMJAU 43709(Holotype!),湘西土家族苗族自治区永顺县小溪国家级自然保护区,娜琴,2018年5月15日,HMJAU 43711,湘西土家族苗族自治区古丈县高望界国家级自然保护区,娜琴,2018年5月17日,HMJAU 43716。
讨论:Singer(1976)提供的采自南美洲哥伦比亚的Mycena heteracantha模式标本的原始描述为:担孢子典型的非淀粉质;褶缘囊状体缺失,所有组织具锁状联合,同时成立了Amparoina,包含了A. spinosissima (Singer) Desjardin和A. heteracantha两个种。随后Desjardin(1995)借阅了Singer的模式标本观察后发现,该种的担孢子强烈淀粉质,褶缘囊状体存在且易发生塌陷,锁状联合未见,因此对该类群重新梳理,强调菌盖表皮和柄囊体权重的基础上提出了全新的分类系统,stirps Amparoina为Mycena sect. Sacchariferae组下等级,A. spinosissima和A. heteracantha作为M. spinosissima和M. heteracantha的异名。Desjardin将采自厄瓜多尔命名为M. asterophora材料订正为M. heteracantha,同时作为M. heteracantha的等模式。
由于M. heteracantha和M. spinosissima的形态学特征极其相似,导致物种界限并不清晰(Horak 1983)。二者菌盖表面均具圆锥状刺突、中央形成麸状或皮屑状绒毛,但M. spinosissima菌盖中央淡黄色,cherocytes紧密排列呈链状,薄壁,较小(Desjardin 1995)。
Mycena hiemalis (Osbeck) Quél., Mém. Soc. Émul. Montbéliard, Sér. 2 5: 110 (1872).
≡Agaricus hiemalis Osbeck, Göteb. Wet. o Witt. Samk. Handl. Afd., styck. 4: 34 (1788).
菌盖直径2.5-9.3mm,钟形、凸镜形,成熟后扁半球形,中央尖突起,褐色、深褐色,向边缘渐浅至米褐色、灰白色,光滑,湿时黏。菌肉白色。无明显气味与味道。菌褶白色,弯生至稍延生,垂齿状,窄。菌柄8-26×0.5-1.0mm,中空,圆柱形,脆骨质,透明状,基部淡黄褐色,表面被细小白色绒毛,基部具白色绒毛。
担孢子6.6-9.7×4.9-7.2μm,Q=1.1-1.5,宽椭圆形,无色,光滑,非淀粉质。担子20-36× 6-8μm,棍棒状,具2个小梗。褶缘囊状体27-52× 8-21μm,棍棒状、纺锤形、泡囊形。褶侧囊状体缺失。菌盖表皮具稀疏瘤疣状或长棒状突起,2.5-8.3×0.6-2.8μm。柄生囊状体15-34×5-12μm,棍棒状至头部膨大,薄壁,表面不平整,呈波浪状。未见锁状联合。
生境:夏季至冬季单生、散生于针阔混交林腐木苔藓层上。
图9
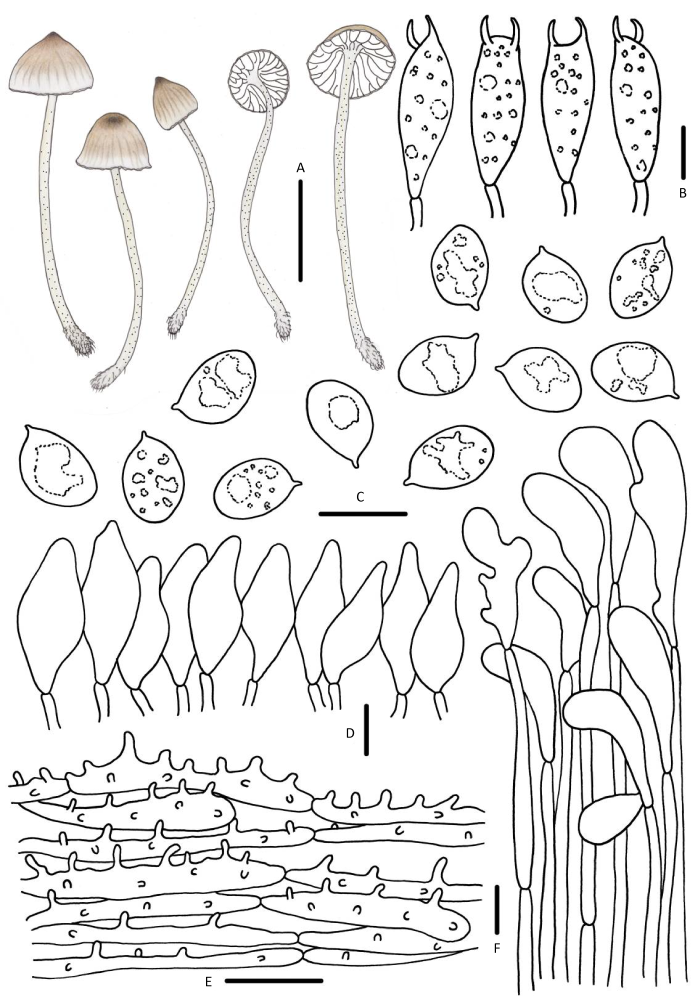
图9
冬生小菇
A:担子体;B:担子;C:担孢子;D:褶缘囊状体;E:菌盖表皮;F:菌柄皮层. 标尺:A=10mm,B-F=10μm
Fig. 9
Mycena hiemalis.
A: Basidiomata; B: Basidia; C: Basidiospores; D: Cheilocystidia; E: Pileipellis; F: Stipitipellis. Bars: A=10mm, B-F=10μm.
中国分布:内蒙古、吉林。
研究标本:内蒙古:呼伦贝尔市满归镇伊克萨玛国家森林公园,娜琴,2017年8月30日,HMJAU 43620。吉林:敦化市胜利河林场,娜琴,2017年6月27日,HMJAU 43512。
讨论:Mycena hiemalis的原始描述为:菌盖钟形,薄,表面具条纹;菌柄细,脆骨质,被粉霜;菌褶弯生至稍延生(Quélet 1872)。
M. alba与M. hiemalis易混淆,但前者担孢子近球形。M. olida与本种相似,但M. olida菌盖白色,中央黄褐色,菌盖表皮菌丝光滑。M. hiemalis在野外工作中易被错误鉴定为M. erubescens,区分特征为后者菌褶灰褐色,老后出现红色斑点,褶缘囊状体具多个树枝状突起(Robich 2003;Aronsen & Læssøe 2016)。
Mycena maculata P. Karst., Meddn Soc. Fauna Flora fenn. 19: 89 (1890).
菌盖直径9-28mm,凸镜形、半球形,后稍平展,中央尖至钝圆突起,浅褐色、褐色、黑褐色,中央红褐色,老后表面形成红褐色斑点,边缘二分之一具半透明状条纹,湿时黏。菌肉白色,薄。气味不明显,味道稍苦涩。菌褶白色,老后褐色,直生,垂齿状,褶间具横脉,老后褶面形成红褐色斑点。菌柄46-92×1.0-3.0mm,上下等粗,中空,光滑,上部灰白色,下部褐色,基部红褐色、深褐色,基部具密集白色绒毛。
担孢子7.5-8.5×4.1-5.0μm,Q=1.3-2.1,椭圆形,无色,光滑,淀粉质。担子24-35×6-7μm,棍棒状,具2或4个小梗。褶缘囊状体棒状,24-46× 10-17μm,头部具不规则瘤突或指状突起。褶侧囊状缺失。菌盖表皮稀疏瘤状突起,1.0-3.0× 0.5-1.0μm。菌柄皮层非光滑,具稀疏瘤疣突。所有组织具锁状联合。
生境:夏秋季群生于云杉、落叶松、红松、槭、栎等针阔混交林腐木上。
中国分布:内蒙古、吉林、黑龙江、四川、云南、西藏。
研究标本:内蒙古:内蒙古呼伦贝尔市满归镇伊克萨玛国家森林公园,娜琴,2016年8月12日,HMJAU 43194,2017年7月30日,HMJAU 43634。吉林:延边朝鲜族自治州安图县二道白河镇科学院后山,娜琴,2016年6月15日,HMJAU 43009,HMJAU 43035,HMJAU 43037,HMJAU 43111;延边朝鲜族自治州安图县三道岭,娜琴,2016年6月17日,HMJAU 43085;敦化市老白山自然保护区,娜琴,2016年8月25日,HMJAU 43236;蛟河市红叶谷,娜琴,2016年8月27日,HMJAU 43264;蛟河市松江镇,娜琴,2017年6月29日,HMJAU 43527;延边朝鲜族自治州安图县二道白河镇老山门,娜琴,2018年9月11日,HMJAU 43845,HMJAU 43846。黑龙江:呼玛县呼中国家级自然保护区,娜琴,2016年8月17日,HMJAU 43224,HMJAU 43225。
讨论:Karsten(1890)首次命名了Mycena maculata,Saccardo(1891)提供了详细的形态学描述:菌盖半球形、钟形,老后平展,中央钝圆突起,红褐色,具红斑,表面条纹;菌柄等粗,光滑,灰白色,基部红褐色,长13cm;菌褶直生,与菌柄连接处锯齿状,成熟后具红斑;担孢子6-9×4-6μm,椭圆形;担子棒状,25-30×6-8μm;生于松树活立木或腐木上,相似种为盔盖小菇Mycena galericulata (Scop.) Gray。
M. maculata在幼时与M. galericulata极为相似,菌盖均为褐色至深褐色,但M. galericulata具强烈的淀粉味,褶缘囊状体细棒状,担孢子较大,宽椭圆形。沟柄小菇M. polygramma (Bull.) Gray菌褶表面在成熟后也具红褐色斑点,但菌柄表面具明显纵条纹,担孢子宽椭圆形,褶缘囊状体光滑或偶见少分枝。另外,M. inclinata (Fr.) Quél.区别与本种的特征为菌盖边缘锯齿状、味道为辛辣味(Robich 2003;Aronsen & Læssøe 2016)。
图10
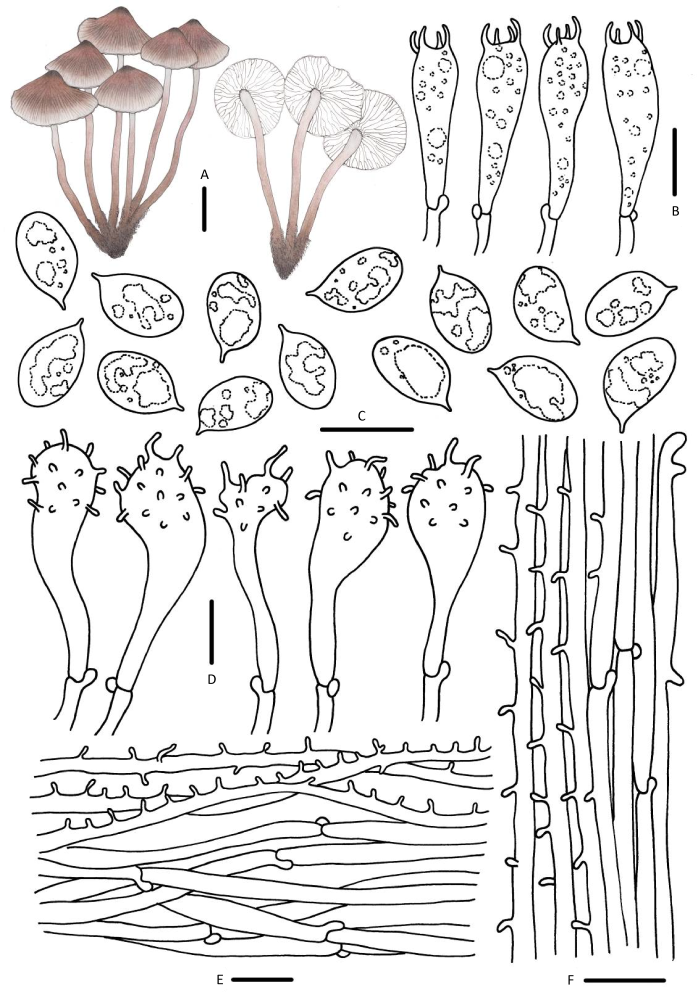
图10
红斑小菇
A:担子体;B:担子;C:担孢子;D:褶缘囊状体;E:菌盖表皮;F:菌柄皮层. 标尺:A=10mm,B-F=10μm
Fig. 10
Mycena maculata.
A: Basidiomata; B: Basidia; C: Basidiospores; D: Cheilocystidia; E: Pileipellis; F: Stipitipellis. Bars: A=10mm, B-F=10μm.
Mycena speirea (Fr.) Gillet, Hyménomycètes (Alençon): 428 (1876) [1878].
≡Agaricus speireus Fr., Observ. mycol. (Havniae) 1: 90 (1815).
≡Phloeomana speirea (Fr.) Redhead, Index Fungorum 15: 2 (2013).
菌盖直径2.5-9mm,凸镜形、半球形,老后扁半球形,中央具乳突,灰褐色至褐色,边缘浅灰褐色、浅褐色,表面被白色细小绒毛,后近光滑,具透明状条纹。菌肉白色。无明显气味与味道。菌褶白色,弯生至稍延生,垂齿状,褶片间不明显横脉。菌柄15-48×0.5-1.5mm,中空,圆柱形,脆骨质,淡黄褐色,下部淡褐色、褐色,光滑,基部具球状白色长纤维状绒毛。
担孢子6.5-9.6×4.2-5.7μm,Q=1.7-1.9,长椭圆形、杏仁形,无色,光滑,非淀粉质。担子22-28×6-8μm,棒状,具2个小梗。褶缘囊状体25-49×5-9μm,拟圆柱形、拟纺锤形、长棒形,表面弯曲呈波浪状。褶侧囊状体缺失。菌盖表皮表面具稀疏瘤疣状或棒状突起,1.2-8.3×0.8-1.5μm。柄生囊状体16-54×4-9μm,圆柱状、棒状,表面呈不规则波浪状。锁状联合未见。
生境:夏秋季单生、散生于壳斗科、华山松等针阔混交林腐木或腐殖质上。
中国分布:吉林、四川、云南。
研究标本:吉林:延边朝鲜族自治州安图县二道白河镇科学院后山,娜琴,2016年6月15日,HMJAU 43107,2017年6月29日,HMJAU 43546,HMJAU 43547;和龙市甄峰岭,娜琴,2016年6月17日,HMJAU 43091;蛟河市前进林场,娜琴,2017年6月26日,HMJAU 43473;蛟河市松江镇,娜琴,2017年6月28日,HMJAU 43520。四川:甘孜藏族自治州捧塔乡水库,娜琴,2017年8月19日,HMJAU 43660。云南:昆明市野鸭湖风景区,娜琴,2017年8月27日,HMJAU 43694。
讨论:M. speirea早期因菌褶延生而划分在亚脐菇属Omphalina中,Kühner(1938)随后修订Mycena属的范围,主张将M. speirea纳入,这一观点也被Smith(1934,1947)认可和采纳。Kühner(1938)观察到M. speirea担子具4个小梗,锁状联合丰富,但本研究的标本及欧洲材料均只观察到2个担子小梗,且未见锁状联合(Robich 2003;Aronsen & Læssøe 2016)。
M. clavata为M. speirea的相近种,但前者菌盖和菌柄咖啡色,担孢子宽椭圆形至近球形,褶缘囊状体纺锤形、梭形(Aronsen & Læssøe 2016)。
图11
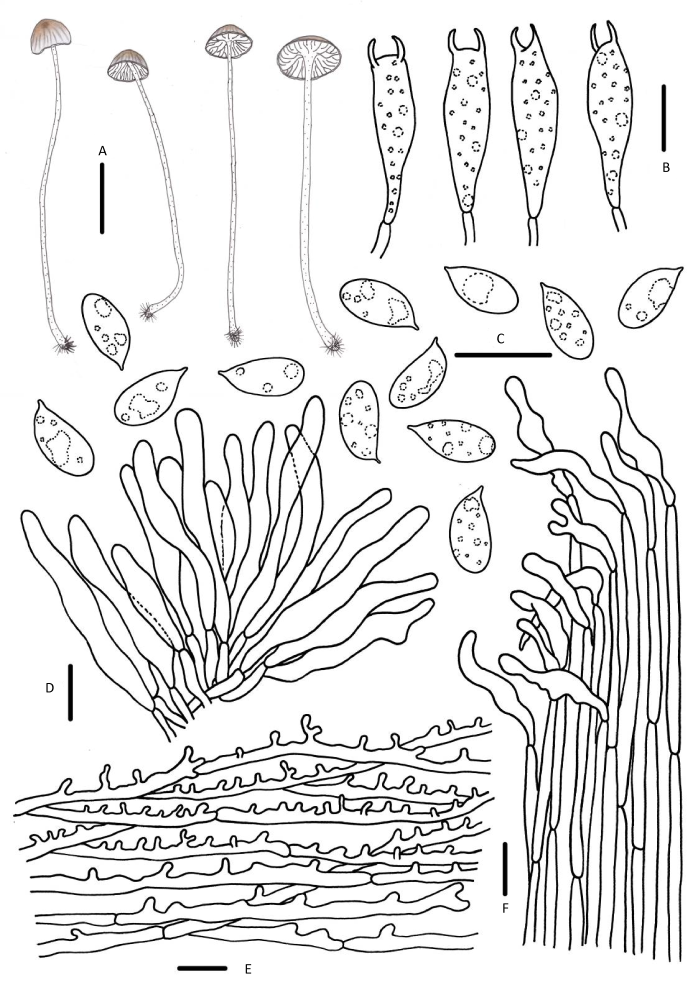
图11
绣线菊小菇
A:担子体;B:担子;C:担孢子;D:褶缘囊状体;E:菌盖表皮;F:菌柄皮层. 标尺:A=10mm,B-F=10μm
Fig. 11
Mycena speirea.
A: Basidiomata; B: Basidia; C: Basidiospores; D: Cheilocystidia; E: Pileipellis; F: Stipitipellis. Bars: A=10mm, B-F=10μm.
Mycena zephirus (Fr.) P. Kumm., Führ. Pilzk. (Zerbst): 110 (1871).
≡Agaricus zephirus Fr., Observ. mycol. (Havniae) 2: 161 (1818).
菌盖直径22-45mm,圆锥形、斗笠形,老后稍平展,中央钝圆突起,淡红褐色、红褐色,中央褐色,边缘乳白色,表面被白色细小绒毛,老后出现红褐色斑点,透明状条纹。菌肉白色。气味与味道淡胡萝卜味。菌褶白色至弯生,垂齿状,老后褶面具红褐色斑点。菌柄34-66×1.5-4.0mm,中空,上下等粗,脆骨质,灰白色,向下渐深至灰褐色,基部褐色,微被白色细小绒毛,基部具丰富白色菌丝体。
担孢子8.1-11.9×4.0-5.5μm,Q=1.9-2.5,长椭圆形、圆柱形,光滑,无色,淀粉质。担子26-34×6-9μm,棍棒状,具4小梗。褶缘囊状体纺锤形、棒状,顶端具几个钝圆分枝,42-68× 5-17μm,光滑。褶侧囊状体缺失。菌盖表皮偶见疣突或圆柱状突起,1.5-6.9×1.0-2.1μm。菌柄皮层具稀疏圆柱状疣突,柄生囊状体23-36× 5-10μm,纺锤形、棍棒状,顶端具钝圆分枝,表面被短突起。具锁状联合。
生境:夏秋季散生于落叶松、红松、云杉等针叶林腐木苔藓层上。
中国分布:辽宁、吉林。
图12
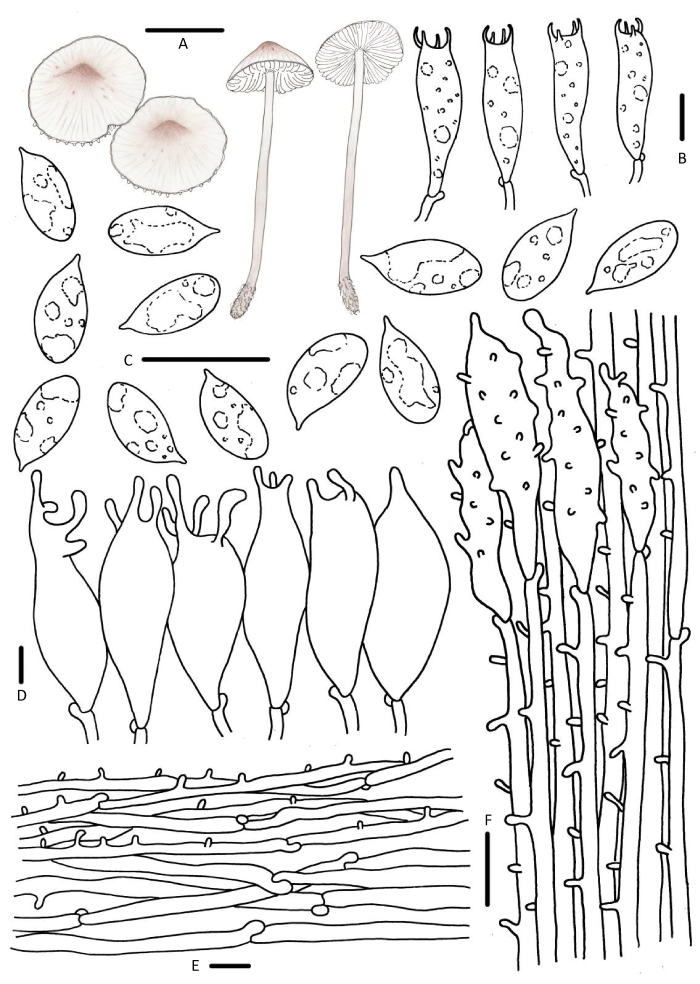
图12
粉被小菇
A:担子体;B:担子;C:担孢子;D:褶缘囊状体;E:菌盖表皮;F:菌柄皮层. 标尺:A=10mm,B-F=10μm
Fig. 12
Mycena zephirus.
A: Basidiomata; B: Basidia; C: Basidiospores; D: Cheilocystidia; E: Pileipellis; F: Stipitipellis. Bars: A=10mm, B-F=10μm.
研究标本:辽宁:丹东市五龙脊镇五龙山,段明政,2016年6月30日,HMJAU 43123。吉林:延边朝鲜族自治州安图县二道白河镇科学院后山,娜琴,2016年6月15日,HMJAU 43028,HMJAU 43036,HMJAU 43106,2018年6月8日,HMJAU 43737;蛟河市杉松岭,娜琴,2016年8月26日,HMJAU 43245;蛟河市红叶谷,娜琴,2016年8月27日,HMJAU 43255。
讨论:菌盖及菌褶老后具红褐色斑点是Mycena zephirus的主要识别特征(Maas Geesteranus 1988)。M. ustalis Aronsen & Maas Geest.与M. zephirus易混淆,但前者褶间具横脉,无红褐色斑点,担孢子较小,7-8.5×5-6μm,褶缘囊状体和柄生囊状体光滑,棍棒状且头部钝圆、纺锤形且顶端渐细(Aronsen & Læssøe 2016)。据报道M. zephirus可以产生荧光且发酵产物提取到抗菌活性成分,为新药开发提供了新线索(Bäuerle et al. 1982;Chew et al. 2015)。
3 讨论
本研究利用经典形态学和分子系统学相结合的方法研究确认了采自多个省份的小菇属10个新记录种。通过与欧洲、北美洲、南美洲、亚洲的材料比较发现,这些物种的形态学特征与之高度吻合,个别种如黄白小菇Mycena flavoalba的担孢子与其他洲的原始描述存在差异。另外,基于我国的材料补充了10个种的生境信息,为更全面地认知该物种提供资料。
基于ITS序列的系统发育分析能够清晰地揭示新记录种的系统位置和系统发育关系,为反映小菇属物种的演化规律提供重要信息。本文记载的10个种分布在小菇属6个组中,随着日后研究材料的不断补充,对物种、组、属的认知将更加深刻。
致谢:感谢湖南师范大学张平教授、长白山科学院王柏先生在野外调查中给予的指导和帮助。感谢海南医学院范宇光博士、鲁东大学盖宇鹏博士和江西农业大学颜俊清博士对本研究的帮助。
参考文献
Mycenas of Kerala.
Antibiotics from Basidiomycetes XVI. Antimicrobial and cytotoxic polyines from Mycena viridimarginata Karst
Studies on the taxonomy of Mycena
Taxonomic studies on Mycena from Guangdong Province of China
Wild edible fungi: a global overview of their use and importance to people. Non-woodforest
Bioluminescent fungi from Peninsular Malaysia-a taxonomic and phylogenetic overview
Six species of Tricholomataceae (Basidiomycotina) new to Taiwan
A revised checklist of medicinal fungi in China
A revised checklist of edible fungi in China
A preliminary accounting of the worldwide members of Mycena sect. Sacchariferae
Les associations fongiques des hauts-marais jurassiens et de quelques regions voisines
Fungal diversity and mechanisms of symbiotic germination of orchid seeds: a review
Mycena anoectochila sp. nov. isolated from mycorrhizal roots of Anoectochilus roxburghii from Xishuangbanna, China
Mycena dendrobii, a new mycorrhizal fungus
Three new species of Mycena
Mycogeography in the South Pacific Region: Agaricales, Boletales
Symbolae ad mycologiam fennicam XXIX
Dictionary of the Fungi. 10th ed
Le genre Mycena (Fries): étude cytologique et systématique des espèces d'Europe et d'Amérique du Nord (Vol. 10)
Studies in Mycena 148-167
Conspectus of the Mycenas of the Northern Hemisphere-9. Section Fragilipedes, species IR
Conspectus of the Mycenas of the northern hemisphere. 14. sections Adonideae, Aciculae, and Oregonenses
Studies in Mycenas. Additions and corrections I
Mycenas of the Northern Hemisphere I. Studies in Mycenas and other papers
Mycenas of the Northern Hemisphere II. Studies in Mycenas and other papers
Major clades of agaricales: a multilocus phylogenetic overview
One hundred and seventeen clades of euagarics
Pleurotus, Omphalia, Mycena and Collybia published in North American Flora
DOI:10.1080/00275514.1916.12018883 URL [本文引用: 1]
Taxonomy and phylogeny of Mycena in China. PhD Dissertation,
New species of Mycena (Mycenaceae, Agaricales) with colored lamellae and three new species records from China
Mycena section Sacchariferae: three new species with basal discs from China
Recognition of Mycena sect. Amparoina sect. nov. (Mycenaceae, Agaricales) including four new species and revision of the limits of sect. Sacchariferae
DOI:10.3897/mycokeys.52.34647
URL
PMID:31148935
[本文引用: 2]

Phylogenetic reconstruction revealed that Mycena stirps Amparoina, which is traditionally classified in sect. Sacchariferae, should be treated at section level. Section Amparoina is characterised by the presence or absence of cherocytes, the presence of acanthocysts and spinulose caulocystidia. Eight species referred to Mycenasect.Amparoinasect. nov. are recognised in China. Of these taxa, four new species classified in the new section are formally described: M.bicystidiata sp. nov., M.griseotincta sp. nov., M.hygrophoroides sp. nov. and M.miscanthi sp. nov. The new species are characterised by the absence of both cherocytes and a basal disc, along with the presence of acanthocysts on the pileus, spinulose cheilocystidia and caulocystidia. Descriptions of the new species, accompanied by illustrations of morphological characters and comparisons with closely related taxa, are provided. A multi-locus analysis utilising the ITS + nLSU + SSU regions was carried out using maximum likelihood and Bayesian Inference. A key to the 12 species of sect. Amparoinasect. nov. and sect. Sacchariferae that are found in China is provided.
Mycenarubins A (I) and B (II), red pyrroloquinoline alkaloids from the mushroom Mycena rosea
MODELTEST: testing the model of DNA substitution
DOI:10.1093/bioinformatics/14.9.817
URL
PMID:9918953
[本文引用: 1]
SUMMARY: The program MODELTEST uses log likelihood scores to establish the model of DNA evolution that best fits the data. AVAILABILITY: The MODELTEST package, including the source code and some documentation is available at http://bioag.byu. edu/zoology/crandall_lab/modeltest.html.
The mushrooms of the Jura and Vosges. Mémoire de la Société d’Emilation de
Mrbayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space
Type studies in basidiomycetes X
Amparoinaceae and Montagneaceae
Investigations of two-spored forms in the genus Mycena
RAxML-III: a fast program for maximum likelyhood-based inference of large phylogenetic trees
DOI:10.1093/bioinformatics/bti191
URL
PMID:15608047
[本文引用: 1]
MOTIVATION: The computation of large phylogenetic trees with statistical models such as maximum likelihood or bayesian inference is computationally extremely intensive. It has repeatedly been demonstrated that these models are able to recover the true tree or a tree which is topologically closer to the true tree more frequently than less elaborate methods such as parsimony or neighbor joining. Due to the combinatorial and computational complexity the size of trees which can be computed on a Biologist's PC workstation within reasonable time is limited to trees containing approximately 100 taxa. RESULTS: In this paper we present the latest release of our program RAxML-III for rapid maximum likelihood-based inference of large evolutionary trees which allows for computation of 1.000-taxon trees in less than 24 hours on a single PC processor. We compare RAxML-III to the currently fastest implementations for maximum likelihood and bayesian inference: PHYML and MrBayes. Whereas RAxML-III performs worse than PHYML and MrBayes on synthetic data it clearly outperforms both programs on all real data alignments used in terms of speed and final likelihood values. Availability SUPPLEMENTARY INFORMATION: RAxML-III including all alignments and final trees mentioned in this paper is freely available as open source code at http://wwwbode.cs.tum/~stamatak CONTACT: stamatak@cs.tum.edu.
The ecological function of wood-inhabiting fungi in forest ecosystem
Wood-inhabiting fungi are one of the most important parts of forest ecosystem, and play an important role in degrading the wood in forest ecosystem. The major species of these fungi include the groups of Aphyllophorales (Basidiomycota), Discomycetes (Ascomycota) and some imperfect fungi. They have the ability to degrade cellulose, hemicelluloses and lignin of wood. Three type of wood decaying have been found, i.e., white rot, brown rot and soft rot. Many other organisms of forest ecosystem have symbiosis relationship with wood-decaying fungi. Wood-inhabiting fungi could offer the nutrition for many insects and birds, and spores of many wood-rotting species are spread by some insects. The high biodiversity of wood-decaying fungi is one of the important factors for the health of forest ecosystem.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}