



木腐真菌并非广义上的分类阶元,而是一类依托倒死木生存的真菌类群的统称,主要包括担子菌门多孔菌目、锈革菌目、子囊菌门盘菌纲、炭角菌科和半知菌类的部分真菌(Jönsson et al. 2008;戴玉成 2009)。它们能较为彻底地降解木材中的木质素和纤维素(Liers et al. 2006),是森林生态系统物质循环中不可或缺的关键部分,其降解残留物有助于林分的自然更新,子实体能为其他生物提供食物与居所(Krajick 2001)。木腐真菌种类繁多,目前我国报道的木腐真菌已超过2 000种(戴玉成和庄剑云 2010;Dai 2010,2011,2012;Cui et al. 2019)。腐生是木腐真菌的主要生活方式,降解模式主要为白腐或褐腐。白腐真菌可分泌一系列的胞外氧化酶,将纤维素和木质素降解为漂白纤维残渣。褐腐真菌则以降解纤维素为主,经分解后的残渣呈立体小方块状(Schwarze 2007;魏玉莲 2021)。林分的优势树种及其产生的倒木数量是影响木腐真菌分布的最主要因素之一,不同种类倒木上木腐真菌种类有很大差异。Krah et al.(2018)发现,与环境条件(即小气候)或寄主大小相比,寄主树种对木腐真菌群落组成和多样性的驱动力更大。Baber et al.(2016)分析了13个树种倒木上的木腐真菌多样性,发现不同林分的不同寄主倒木上真菌物种丰富度和群落组成均有很大差异,其主要原因在于树种木材理化性质的不同影响了木腐真菌种类的选择和生长。除了寄主倒木种类的影响外,木腐真菌的分布还受诸多因素影响,包括经纬度、年平均降水量、年均温、海拔等。Tedersoo et al.(2014)的研究表明,距赤道的距离和年平均降水量对真菌丰富度的影响最大,包括大多数真菌的分类单元和功能群,多数真菌类群多样性在热带森林生态系统中达到高峰。
我国有较大范围热带雨林覆盖的区域,主要分布在海南省与云南省西双版纳地区。两区域以重要值排序树种的优势科存在较大重叠,组成接近,有基本一致的植物区系组成。海南地区热带雨林类型依照其所处的海拔位置,划分为热带低地雨林和热带山地雨林,而热带季节性雨林则是西双版纳地区热带雨林的主要植被类型(朱华和周虹霞 2002)。但两个地区在树种组成方面存在明显差异,以龙脑香热带雨林为例,西双版纳的优势种为假海桐Pittosporopsis kerrii和望天树Shorea wantianshuea,海南则为青梅Vatica mangachapoi(兰国玉等 2010)。吴芳等(2020)系统地报道了我国华南地区的多孔菌500种,在华南地区发展多孔菌新属10个、新种132个。戴玉成和崔宝凯(2010)对海南地区大型木腐真菌进行全面考察的基础上,报道了240种木生真菌,它们以泛热带成分和北温带成分为主,表现出明显的泛热带特征,其中93%左右为白腐真菌(员瑗等 2015)。而同属于热带地区的西双版纳,目前还未见有关木腐真菌物种组成和分布特征的相关文献,仅有新种的发表和报道。那么我国热带地区不同林分中的木腐真菌物种组成是否相似?如有分布差异,主要影响因子是什么?
为探究我国热带地区木腐真菌的物种组成及其分布特征,本研究选取海南尖峰岭、霸王岭及云南西双版纳为代表性研究地点,基于十多年的标本采集数据,统计木腐真菌物种组成及其生态习性,并进行比较,查明我国热带地区木腐真菌群落组成,为保护热带地区木腐真菌资源起积极推动作用。
1 材料与方法
1.1 研究区域概况
1.1.1 尖峰岭国家级自然保护区:位于海南岛西南部,地跨乐东和东方两县市(18°20ʹ -18°57ʹ N,108°41ʹ-109°12ʹ E),属低纬度热带岛屿季风气候,干湿两季明显,年均温24.5℃,年均降水量介于1 300-3 700mm。热带山地雨林为该地区面积最大的植被类型,主要优势树种有:大叶蒲葵Livistona saribus、白颜树Gironniera subaequalis、厚壳桂Cryptocarya chinensis、油丹Alseodaphne hainanensis、三角瓣花Prismatomeris tetrandra、海南韶子Nephelium topengii、红柯Lithocarpus fenzelianus、香果新木姜子Neolitsea ellipsoidea、东方琼南Beilschmiedia tungfangensis、九节Psychotria asiatica、卵叶桂Cinnamomum rigidissimum(许涵等 2015)。
1.1.2 霸王岭国家级自然保护区:隶属于海南省昌江县,位于白沙、昌江县之间(18°50ʹ- 19°05ʹ N,109°05ʹ-109°25ʹ E),属热带岛屿季风气候,干湿两季明显,年均温23.6℃,年均降水量介于1 500-2 000mm。主要优势树种有:线枝蒲桃Syzygium araiocladum、三角瓣花、灰木Symplocos spp.、粗叶木Lasianthus chinensis、谷木Memecylon spp.、九节、陆均松Dacrydium pierrei、卵叶桂、赤营Syzygium buxifolium、厚壳桂(臧润国等 2001;吕晓波等 2012;刘浩栋等 2020)。
1.1.3 西双版纳国家级自然保护区:位于云南南部的西双版纳傣族自治州境内(21°08ʹ- 22°36ʹ N,99°56ʹ-101°50ʹ E),与老挝接壤,毗邻缅甸,属热带季风气候,干湿两季明显,年均温21.8℃,年均降水量为1 493mm,海拔范围从澜沧江河谷最低点(491m)至最高峰(2 492m)。热带季节雨林和热带山地常绿阔叶林为该地区的主要植被类型,主要优势树种有:假海桐、望天树、云树Garcinia cowa、短刺锥Castanopsis echidnocarpa、蚁花Mezzettiopsis creaghii、毛猴欢喜Sloanea tomentosa、木奶果Baccaurea ramiflora、红光树Knema furfuracea、绒毛番龙眼Pometia tomentosa、披针叶楠Phoebe lanceolata(兰国玉等 2008)。
1.2 研究资料来源与处理
本研究主要基于近十多年来东北生物标本馆和北京林业大学生态与自然保护学院微生物研究所对海南尖峰岭、霸王岭和云南西双版纳地区的木腐真菌开展的资源调查与标本采集工作。对各地点的木腐真菌种类分别进行了整理,在此基础上对比了各地点已发现或已记录的物种组成。名录中的真菌物种名称及其所属的分类阶元依据Index Fungorum网站(
2 结果与分析
2.1 我国热带地区木腐真菌物种组成
表1 海南尖峰岭、霸王岭及云南西双版纳木腐真菌名录
Table 1
| 种名 Species | 尖峰岭 Jianfeng- ling | 霸王岭 Bawang- ling | 西双版纳 Xishuang- banna | 出现频率 Occurrence | 腐朽类型 Decay types |
|---|---|---|---|---|---|
| Aleurodiscus botryosus | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Aleurodiscus tenuissimus | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Aleurodiscus thailandicus | - | + | - | 常见 Common | 白色腐朽 White-rot |
| Amauroderma preussii | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Amauroderma scopulosum | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Amylosporia hattorii | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Aporpium efibulatum | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Asterostroma muscicola | + | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Auricularia cornea | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Auricularia delicata | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Auricularia fibrillifera | + | + | + | 偶见 Occasional | 白色腐朽 White-rot |
| Auricularia heimuer | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Auricularia nigricans | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Auricularia orientalis | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Baltazaria galactina | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Bjerkandera adusta | + | - | + | 常见 Common | 白色腐朽 White-rot |
| Bondarzewia podocarpi | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Bresadolia cuticulata | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Butyrea luteoalba | - | - | + | 常见 Common | 白色腐朽 White-rot |
| Byssomerulius corium | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Calyptella capula | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Campanella junghuhnii | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Castanoporus castaneus | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Cerioporus mollis | - | + | - | 常见 Common | 白色腐朽 White-rot |
| Ceriporia lacerata | - | + | + | 常见 Common | 白色腐朽 White-rot |
| Ceriporia mellita | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Ceriporia spissa | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Ceriporia tarda | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Ceriporia viridans | + | - | - | 常见 Common | 白色腐朽 White-rot |
| Ceriporiopsis albonigrescens | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Ceriporiopsis carnegieae | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Ceriporiopsis mucida | + | - | - | 常见 Common | 白色腐朽 White-rot |
| Cerrena unicolor | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Cerrena zonata | + | - | + | 常见 Common | 白色腐朽 White-rot |
| Climacodon pulcherrimus | + | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Coltriciella dependens | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Coltriciella subglobosa | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Coltriciella tasmanica | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Coriolopsis brunneoleuca | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Coriolopsis gallica | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Crassisporus imbricatus | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Crustodontia chrysocreas | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Cryptoporus sinensis | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Cyclomyces setiporus | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Cymatoderma elegans | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Dacryobolus gracilis | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Daedalea radiata | - | - | + | 偶见 Occasional | 褐色腐朽 Brown-rot |
| Daedaleopsis nipponica | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Dentipellicula taiwaniana | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Dentipellis tropicalis | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Dichostereum pallescens | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Duportella tristicula | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Earliella scabrosa | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Echinoporia hydnophora | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Elmerina cladophora | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Epithele malaiensis | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Erastia salmonicolor | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Erythromyces crocicreas | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Favolaschia pustulosa | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Favolus acervatus | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Favolus emerici | + | + | + | 偶见 Occasional | 白色腐朽 White-rot |
| Favolus philippinensis | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Favolus pseudoemerici | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Favolus spathulatus | - | + | + | 偶见 Occasional | 白色腐朽 White-rot |
| Fistulina subhepatica | - | + | - | 稀有 Rare | 褐色腐朽 Brown-rot |
| Flabellophora superposita | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Flaviporus liebmannii | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Flaviporus stramineus | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Flavodon flavus | - | + | + | 常见 Common | 白色腐朽 White-rot |
| Fomitiporella coruscans | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Fomitiporella sinica | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Fomitiporia bannaensis | + | - | + | 常见 Common | 白色腐朽 White-rot |
| Fomitiporia hainaniana | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Fomitiporia pseudopunctata | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Fomitiporia robusta | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Fomitopsis massoniana | - | + | - | 稀有 Rare | 褐色腐朽 Brown-rot |
| Fomitopsis palustris | - | + | - | 常见 Common | 褐色腐朽 Brown-rot |
| Fulvifomes durissimus | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Fulvifomes fastuosus | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Fulvifomes glaucescens | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Fulvifomes indicus | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Fulvifomes inermis | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Fulvifomes merrillii | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Fulvifomes minisporus | - | + | + | 常见 Common | 白色腐朽 White-rot |
| Fulvifomes rimosus | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Funalia aspera | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Funalia sanguinaria | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Fuscoporia bambusicola | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Fuscoporia chrysea | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Fuscoporia discipes | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Fuscoporia ferrea | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Fuscoporia gilva | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Fomitiporia pusilla | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Fuscoporia rhabarbarina | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Fuscoporia senex | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Fuscoporia torulosa | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Fuscoporia undulata | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Fuscoporia wahlbergii | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Ganoderma australe | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Ganoderma calidophilum | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Ganoderma flexipes | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Ganoderma gibbosum | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Ganoderma hoehnelianum | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Ganoderma lobatum | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Ganoderma multiplicatum | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Ganoderma nigrolucidum | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Ganoderma orbiforme | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Ganoderma philippii | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Ganoderma shangsiense | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Ganoderma sinense | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Ganoderma subresinosum | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Ganoderma tropicum | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Ganoderma weberianum | + | - | - | 常见 Common | 白色腐朽 White-rot |
| Gloeodontia eriobotryae | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Gloeophyllum striatum | - | + | - | 稀有 Rare | 褐色腐朽 Brown-rot |
| Gloeoporus hainanensis | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Grammothele fuligo | + | - | - | 常见 Common | 白色腐朽 White-rot |
| Grammothele lineata | - | + | + | 偶见 Occasional | 白色腐朽 White-rot |
| Grammothele subargentea | - | - | + | 常见 Common | 白色腐朽 White-rot |
| Grammothelopsis asiatica | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Grammothelopsis bambusicola | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Gyrodontium sacchari | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Hapalopilus flavus | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Haploporus alabamae | - | + | - | 常见 Common | 白色腐朽 White-rot |
| Henningsomyces subiculatus | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Heterobasidion insulare | + | - | - | 常见 Common | 白色腐朽 White-rot |
| Heterochaete delicata | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Heterochaete roseola | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Heterochaete sinensis | - | + | + | 偶见 Occasional | 白色腐朽 White-rot |
| Hexagonia cucullata | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hexagonia glabra | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Hispidaedalea imponens | + | - | - | 偶见 Occasional | 褐色腐朽 Brown-rot |
| Hornodermoporus latissimus | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Humphreya coffeata | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Hydnoporia corrugata | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Hydnoporia latesetosa | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Hymenochaete adusta | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Hymenochaete anomala | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Hymenochaete asetosa | - | + | - | 常见 Common | 白色腐朽 White-rot |
| Hymenochaete attenuata | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete cinnamomea | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete cyclolamellata | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete duportii | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete epichlora | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete floridea | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete innexa | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete legeri | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete lictor | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete longispora | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Hymenochaete luteobadia | + | + | + | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete minor | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete murina | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Hymenochaete nanospora | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete ochromarginata | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete rhabarbarina | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete rheicolor | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Hymenochaete rubiginosa | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete separabilis | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete spathulata | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete sphaerospora | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hymenochaete tropica | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Hymenochaete villosa | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Hymenochaete yunnanensis | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Hyphoderma ayresii | - | + | + | 偶见 Occasional | 白色腐朽 White-rot |
| Hyphodermella poroides | + | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Hyphodontia arguta | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Hyphodontia subglobosa | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Hyphodontia tropica | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Inonotus chrysomarginatus | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Inonotus hainanensis | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Inonotus latemarginatus | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Inonotus lloydii | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Inonotus niveomarginatus | - | + | + | 稀有 Rare | 白色腐朽 White-rot |
| Inonotus pachyphloeus | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Inonotus rigidus | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Inonotus tabacinus | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Inonotus tricolor | + | + | + | 稀有 Rare | 白色腐朽 White-rot |
| Irpex lacteus | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Irpex vellereus | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Kneiffiella abdita | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Laetiporus versisporus | + | - | - | 偶见 Occasional | 褐色腐朽 Brown-rot |
| Lamelloporus americanus | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Larssoniporia tropicalis | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Lentinus arcularius | - | + | - | 常见 Common | 白色腐朽 White-rot |
| Lentinus sajor-caju | + | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Lentinus squarrosulus | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Lenzites acuta | - | + | + | 偶见 Occasional | 白色腐朽 White-rot |
| Leucophellinus hobsonii | - | + | + | 常见 Common | 白色腐朽 White-rot |
| Lindtneria flava | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Lindtneria trachyspora | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Lopharia cinerascens | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Megasporia ellipsoidea | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Megasporia hexagonoides | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Megasporia major | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Megasporia tropica | + | - | - | 常见 Common | 白色腐朽 White-rot |
| Megasporia violacea | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Megasporoporia bannaensis | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Megasporoporia cystidiolophora | + | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Megasporoporia minuta | + | + | + | 偶见 Occasional | 白色腐朽 White-rot |
| Megasporoporia setulosa | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Meiorganum curtisii | - | + | - | 偶见 Occasional | 褐色腐朽 Brown-rot |
| Melanoderma microcarpum | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Mensularia radiata | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Meruliopsis crassitunicata | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Microporus affinis | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Microporus xanthopus | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Murinicarpus subadustus | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Neofavolus alveolaris | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Neofavolus mikawae | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Neofomitella fumosipora | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Neofomitella rhodophaea | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Neomensularia kanehirae | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Nigrofomes melanoporus | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Nigrofomes sinomelanoporus | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Nigroporus durus | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Nigroporus vinosus | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Niveoporofomes spraguei | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Oxyporus corticola | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Oxyporus ginkgonis | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Oxyporus subulatus | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Panellus pusillus | + | - | - | 常见 Common | 白色腐朽 White-rot |
| Panus neostrigosus | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Parmastomyces corticola | + | - | - | 常见 Common | 褐色腐朽 Brown-rot |
| Parmastomyces mollissimus | + | - | - | 常见 Common | 褐色腐朽 Brown-rot |
| Peniophora incarnata | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Peniophora malaiensis | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Perenniporia afrominuta | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Perenniporia bannaensis | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Perenniporia cinereofusca | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Perenniporia cystidiata | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Perenniporia gomezii | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Perenniporia medulla-panis | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Perenniporia subacida | - | + | - | 常见 Common | 白色腐朽 White-rot |
| Perenniporia substraminea | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Perenniporia subtephropora | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Phaeolus schweinitzii | - | + | - | 偶见 Occasional | 褐色腐朽 Brown-rot |
| Phanerochaete australis | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Phellinidium lamaënse | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Phellinus adamantinus | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Phellinus austrosinensis | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Phellinus ellipsoideus | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Phellinus tenuiculus | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Phellopilus nigrolimitatus | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Phlebia subserialis | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Phlebia tremellosa | + | - | - | 常见 Common | 白色腐朽 White-rot |
| Phylloporia clausenae | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Phylloporia dependens | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Phylloporia hainaniana | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Phylloporia minutipora | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Phylloporia pectinata | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Phylloporia pendula | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Phylloporia pseudopectinata | + | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Phylloporia pulla | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Phylloporia ribis | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Phylloporia splendida | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Physisporinus lavendulus | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Picipes badius | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Pilatoporus subtropicus | + | - | - | 稀有 Rare | 褐色腐朽 Brown-rot |
| Piptoporellus hainanensis | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Pleurotus giganteus | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Pleurotus ostreatus | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Polyporus dictyopus | - | + | - | 常见 Common | 白色腐朽 White-rot |
| Polyporus leprieurii | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Polyporus minor | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Polyporus rugulosus | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Polyporus trametoides | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Polyporus tuberaster | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Poria hypobrunnea | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Porpomyces submucidus | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Postia alni | + | + | - | 常见 Common | 褐色腐朽 Brown-rot |
| Postia caesia | - | + | - | 常见 Common | 褐色腐朽 Brown-rot |
| Postia lactea | + | - | - | 稀有 Rare | 褐色腐朽 Brown-rot |
| Pseudofavolus tenuis | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Pseudolagarobasidium acaciicola | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Pseudolagarobasidium calcareum | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Pycnoporus sanguineus | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Pyrofomes castanopsidis | - | + | - | 常见 Common | 白色腐朽 White-rot |
| Pyrrhoderma hainanense | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Pyrrhoderma lamaoense | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Pyrrhoderma noxium | + | - | + | 常见 Common | 白色腐朽 White-rot |
| Pyrrhoderma yunnanense | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Radulotubus resupinatus | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Rajchenbergia tenuissima | - | - | + | 常见 Common | 白色腐朽 White-rot |
| Ranadivia modesta | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Resinicium confertum | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Resiniporus resinascens | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Resinoporia pinea | - | + | - | 常见 Common | 褐色腐朽 Brown-rot |
| Resinoporia sordida | - | + | - | 偶见 Occasional | 褐色腐朽 Brown-rot |
| Rhodofomes subfeei | + | - | - | 常见 Common | 褐色腐朽 Brown-rot |
| Rhodofomitopsis feei | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Rigidoporus microporus | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Rigidoporus minutus | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Rigidoporus ulmarius | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Rigidoporus vinctus | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Royoporus spatulatus | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Rubellofomes cystidiatus | - | + | - | 偶见 Occasional | 褐色腐朽 Brown-rot |
| Sanghuangporus zonatus | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Sanguinoderma elmerianum | + | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Sanguinoderma rugosum | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Schizophyllum commune | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Scytinostroma duriusculum | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Serpula similis | + | + | - | 常见 Common | 褐色腐朽 Brown-rot |
| Sidera vulgaris | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Skeletocutis albomarginata | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Skeletocutis alutacea | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Skeletocutis carneogrisea | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Skeletocutis luteolus | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Skeletocutis nivea | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Skeletocutis stellae | - | + | - | 常见 Common | 白色腐朽 White-rot |
| Skeletocutis substellae | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Skeletocutis subvulgaris | - | + | - | 常见 Common | 白色腐朽 White-rot |
| Sparsitubus nelumbiformis | + | + | + | 偶见 Occasional | 白色腐朽 White-rot |
| Stecchericium seriatum | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Steccherinum fimbriatum | + | - | - | 常见 Common | 白色腐朽 White-rot |
| Steccherinum ochraceum | - | + | - | 常见 Common | 白色腐朽 White-rot |
| Stereum durum | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Stereum gausapatum | - | + | + | 常见 Common | 白色腐朽 White-rot |
| Stereum ostrea | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Theleporus calcicolor | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Theleporus membranaceus | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Theleporus minisporus | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Tinctoporellus epimiltinus | + | - | + | 常见 Common | 白色腐朽 White-rot |
| Trametes apiaria | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Trametes elegans | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Trametes glabrorigens | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Trametes hirsuta | - | + | + | 常见 Common | 白色腐朽 White-rot |
| Trametes orientalis | - | + | + | 偶见 Occasional | 白色腐朽 White-rot |
| Trametes polyzona | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Trametes vernicipes | + | - | + | 常见 Common | 白色腐朽 White-rot |
| Trametes versicolor | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Trametes vespacea | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Trametes villosa | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Trechispora nivea | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Tremella fuciformis | - | - | + | 稀有 Rare | 白色腐朽 White-rot |
| Trichaptum abietinum | - | + | - | 常见 Common | 白色腐朽 White-rot |
| Trichaptum brastagii | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Trichaptum byssogenum | - | + | + | 偶见 Occasional | 白色腐朽 White-rot |
| Trichaptum imbricatum | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Trichaptum podocarpi | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Trullella duracina | + | + | + | 偶见 Occasional | 白色腐朽 White-rot |
| Truncospora fuscopurpurea | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Truncospora ochroleuca | - | - | + | 常见 Common | 白色腐朽 White-rot |
| Truncospora roseoalba | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Truncospora tephropora | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Tyromyces chioneus | + | - | - | 常见 Common | 白色腐朽 White-rot |
| Ungulidaedalea fragilis | + | - | - | 偶见 Occasional | 褐色腐朽 Brown-rot |
| Veluticeps microspora | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Vitreoporus dichrous | + | - | - | 常见 Common | 白色腐朽 White-rot |
| Wolfiporia pseudococos | + | - | - | 稀有 Rare | 褐色腐朽 Brown-rot |
| Wrightoporia austrosinensis | - | + | - | 稀有 Rare | 白色腐朽 White-rot |
| Wrightoporia avellanea | - | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Wrightoporia luteola | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Wrightoporia subavellanea | + | - | - | 偶见 Occasional | 白色腐朽 White-rot |
| Wrightoporiopsis amylohypha | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Wrightoporiopsis irregularis | + | - | - | 稀有 Rare | 白色腐朽 White-rot |
| Xylobolus annosus | + | + | - | 常见 Common | 白色腐朽 White-rot |
| Xylobolus frustulatus | - | + | - | 常见 Common | 白色腐朽 White-rot |
| Xylobolus princeps | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
| Xylobolus spectabilis | + | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Xylodon flaviporus | + | + | + | 常见 Common | 白色腐朽 White-rot |
| Xylodon hallenbergii | - | - | + | 偶见 Occasional | 白色腐朽 White-rot |
| Yuchengia narymica | + | + | - | 偶见 Occasional | 白色腐朽 White-rot |
注:“+”表示此种在该地区发现;“-”表示未发现
Note: “+” indicates the species was collected in the area; “-” indicates that was not collected.
图1
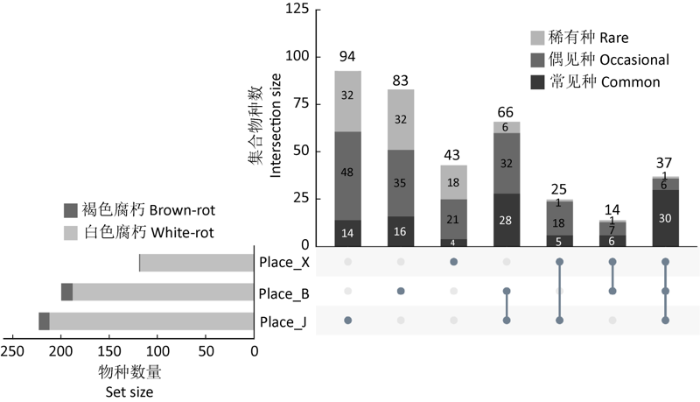
图1
不同区域种分布UpSet图 Place_J:尖峰岭;Place_B:霸王岭;Place_X:西双版纳. 不同区域点线连接情况代表交集类别
Fig. 1
UpSet plot of species distribution in different areas investigated. Place_J: Jianfengling; Place_B: Bawangling; Place_X: Xishuangbanna. The connection of points and line in different areas represents the intersection category.
在本次统计结果中,只出现在尖峰岭的有94种,霸王岭有83种,西双版纳有43种,均以偶见种、稀有种为主,分别占各自种类数的85.11%、80.72%、90.70%;三地区木腐真菌物种组成差异很大。尖峰岭、霸王岭共有种66种,尖峰岭、西双版纳共有种25种,霸王岭、西双版纳共有种14种;三地区共有种为37种,主要为常见种,占81.08%(图1)。
2.2 优势科属情况
三地区木腐真菌的优势科为多孔菌科和锈革菌科。其中尖峰岭多孔菌科包含32属63种,锈革菌科包含17属68种,分别占该地区总种数的28.38%和30.63%;霸王岭地区多孔菌科包含37属69种,占该地区的34.50%,锈革菌科包含9属46种,占该地区的23.00%;西双版纳地区多孔菌科包含21属38种,占该地区的31.93%,锈革菌科包含15属32种,占该地区的26.89%。
尖峰岭地区前3优势属分别为锈革菌属Hymenochaete,22种,占该地总种数的9.91%;灵芝属Ganoderma,11种,占4.95%;褐卧孔菌属Fuscoporia,8种,占3.60%。霸王岭地区前3优势属分别为锈革菌属,有20种,占该地总种数的10.00%;灵芝属,有10种,占5.00%;褐卧孔菌属,有7种,占3.50%。西双版纳地区的优势属并不明显,大多数属的种类数比较接近,相比而言栓孔菌属、锈革菌属、纤孔菌属Inonotus、木耳属Auricularia、褐卧孔菌属、灵芝属、硬孔菌属Rigidoporus、截孢孔菌属Truncospora的种类数略多(表2)。
表2 三地区的优势属种组成情况
Table 2
| 研究区域 Study area | 科名 Family | 属名 Genus | 种数 Numbers of species | 占比 Percentage (%) |
|---|---|---|---|---|
| 尖峰岭 Jianfengling | 木耳科 Auriculariaceae | 木耳属 Auricularia | 5 | 2.25 |
| 锈革菌科 Hymenochaetaceae | 锈革菌属 Hymenochaete 褐卧孔菌属 Fuscoporia 黄层孔菌属 Fulvifomes 叶孔菌属 Phylloporia 嗜蓝孢孔菌属 Fomitiporia 纤孔菌属 Inonotus | 22 8 6 6 5 5 | 9.91 3.60 2.70 2.70 2.25 2.25 | |
| 结晶伏孔菌科 Incrustoporiaceae | 干皮孔菌属 Skeletocutis | 5 | 2.25 | |
| 多孔菌科 Polyporaceae | 灵芝属 Ganoderma 巨孔菌属Megasporia 多年卧孔菌属 Perenniporia | 11 5 5 | 4.95 2.25 2.25 | |
| 霸王岭 Bawangling | 木耳科 Auriculariaceae | 木耳属 Auricularia | 5 | 2.50 |
| 锈革菌科 Hymenochaetaceae | 锈革菌属 Hymenochaete 褐卧孔菌属 Fuscoporia 纤孔菌属 Inonotus 叶孔菌属 Phylloporia | 20 7 5 5 | 10.00 3.50 2.50 2.50 | |
| 结晶伏孔菌科 Incrustoporiaceae | 干皮孔菌属 Skeletocutis | 6 | 3.00 | |
| 多孔菌科 Polyporaceae | 灵芝属 Ganoderma 栓孔菌属 Trametes 巨孔菌属Megasporia | 10 6 4 | 5.00 3.00 2.00 | |
| 西双版纳 Xishuangbanna | 木耳科 Auriculariaceae | 木耳属 Auricularia 刺皮属Heterochaete | 4 3 | 3.36 2.52 |
| 锈革菌科 Hymenochaetaceae | 锈革菌属 Hymenochaete 纤孔菌属 Inonotus 褐卧孔菌属 Fuscoporia 叶孔菌属 Phylloporia 红皮孔菌属 Pyrrhoderma | 5 5 4 3 3 | 4.20 4.20 3.36 2.52 2.52 | |
| 巨盖孔菌科 Meripilaceae | 硬孔菌属 Rigidoporus | 4 | 3.36 | |
| 多孔菌科 Polyporaceae | 栓孔菌属 Trametes 灵芝属 Ganoderma 截孢孔菌属 Truncospora 棱孔菌属 Favolus | 6 4 4 3 | 5.04 3.36 3.36 2.52 |
2.3 热带地区木腐真菌的生态习性
3个研究地点的木腐真菌中白腐真菌有340种,占93.92%,褐腐真菌22种,仅占6.08%。尖峰岭地区的褐腐真菌有11种:Hispidaedalea imponens、Laetiporus versisporus、Parmastomyces corticola、P. mollissimus、Pilatoporus subtropicus、Postia alni、P. lactea、Rhodofomes subfeei、Serpula similis、Ungulidaedalea fragilis、Wolfiporia pseudococos;霸王岭地区的褐腐真菌有12种:Fistulina subhepatica、Fomitopsis palustris、F. massoniana、Gloeophyllum striatum、Meiorganum curtisii、Phaeolus schweinitzii、Postia alni、P. caesia、Resinoporia pinea、R. sordida、Rubellofomes cystidiatus、Serpula similis;西双版纳地区的褐腐真菌有1种:Daedalea radiata。尖峰岭、霸王岭的褐腐真菌占比基本一致,分别为4.95%和6.00%;西双版纳仅0.84%(图1)。
3个研究地点有337种木腐真菌采集于阔叶树倒木上,占93.09%,仅15种采集于针叶树倒木上,占4.14%。尖峰岭有211种采集于阔叶树倒木上,白腐真菌占95.73%,5种于针叶树倒木上获得,含褐腐真菌1种Postia lactea;霸王岭有185种采集于阔叶树倒木上,白腐真菌占97.29%,11种于针叶树倒木上获得,含6种褐腐真菌:Fomitopsis massoniana、Meiorganum curtisii、Phaeolus schweinitzii、Postia caesia、Resinoporia pinea、R. sordida;西双版纳有118种全部采集于阔叶树倒木上,其中白腐真菌占99.15%(图2)。
图2
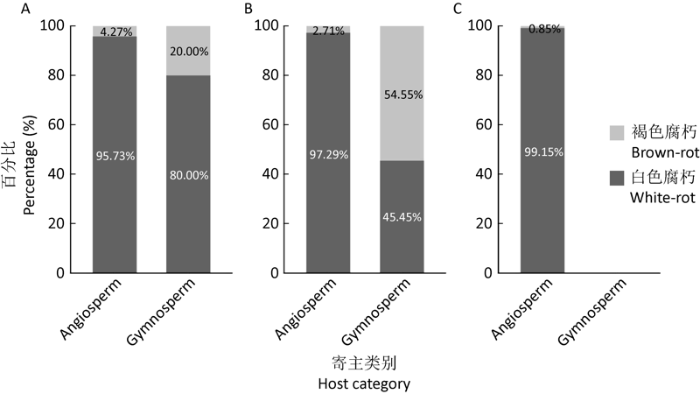
图2
不同地区木腐真菌寄主类别占比情况 A:尖峰岭;B:霸王岭;C:西双版纳
Fig. 2
The proportion of host category of wood-decaying fungi in different regions. A: Jianfengling; B: Bawangling; C: Xishuangbanna.
3 讨论
本研究结果显示,尽管同属于热带地区,三地区362种木腐真菌中仅有37个共有种,且多数为常见种,包括Auricularia delicata、Cyclomyces setiporus、Earliella scabrosa、Hymenochaete luteobadia、Inonotus tabacinus、Megasporoporia minuta、Nigroporus durus、Pseudofavolus tenuis、Skeletocutis albomarginata、Sparsitubus nelumbiformis、Rigidoporus minutus等;稀有种仅1种,为Inonotus tricolor;Megasporoporia minuta为中国特有。这些物种在热带地区极为常见且主要生长于热带地区,可视为热带地区的标志物种。有些物种则是全国广泛分布种,包括Fuscoporia gilva、Hydnoporia corrugata、Lopharia cinerascens、Pycnoporus sanguineus、Xylodon flaviporus,还有一些物种广泛分布于我国亚热带和热带地区,如Auricularia cornea、Ganoderma australe、Hymenochaete anomala、Microporus affinis、M. xanthopus、Rigidoporus microporus、R. vinctus、Stereum ostrea、Truncospora tephropora。实际上,研究结果中的各地特有种,主要是指在本次比较研究中仅出现在该地区的物种,这些物种在其他地区也有分布,可以看出,热带地区的木腐真菌种类既有适应本地气候的特有种,也与其他气候带的物种保持连续性和共通性。出现这种情况的主要原因推测与各地点林分中树种差异有关。目前为止世界范围内只在这3个地区发现的特有种极少,主要为报道于该地区的新种,如Favolus pseudoemerici、Fomitiporella sinica等(Zhou & Cui 2017)。
在中国,仅海南、西双版纳地区,存在真正意义上的热带雨林,热带地区植物区系的科属组成具较高的相似性,分别在90%和64%以上,但树种相似性低于50%(朱华 2017)。三研究地点的年均温、降水情况、气候类型相近。从树种组成来看,尖峰岭和霸王岭具有共同的优势树种:厚壳桂、三角瓣花、九节、卵叶桂等。同海南地区相比,西双版纳的优势树种完全不同。诸多研究表明,林分中的优势树种是木腐真菌群落最主要的影响因素之一,不同种类倒木上的真菌组成存在很大差异(Cui & Dai 2012;王小燕和魏玉莲 2015;李通等 2017;李俊凝等 2020)。相比而言,尖峰岭、霸王岭物种组成相似度较高,共有种66个,而西双版纳则与上述两地存在较大差异,尖峰岭、西双版纳共有种25个,霸王岭、西双版纳的共有种仅14个,这与林分树种的组成差异大体一致。
木腐真菌的寄主分布具有非常鲜明的特点,崔宝凯和余长军(2011)分析了大兴安岭地区多孔菌的区系组成和种群结构,发现白腐真菌更倾向生长在阔叶树倒木上,褐腐真菌则主要生长在针叶树倒木上;整个东北地区多孔类木腐菌中的褐腐真菌所占比例远高于世界平均水平(李敬文和魏玉莲 2014);王小燕和魏玉莲(2015)比较分析了我国西北和东南地区的多孔类木腐真菌物种组成,发现由北至南白腐菌所占比例呈上升趋势,褐腐菌呈下降趋势,其主要原因是因为植被类群中针叶树逐渐减少,而阔叶树大幅增多。本研究结果显示,我国热带地区的木腐真菌中绝大多数为白腐类型(边禄森和戴玉成 2015;武英达等 2021),其主要原因依然是树种组成的差异,霸王岭有陆均松的分布,尖峰岭则生长有竹叶松Podocarpus neriifolius(许涵等 2015),针叶树的存在使得这两个地点的褐腐真菌数量高于西双版纳。本研究结果是基于已有采样数据所开展的寄主与木腐真菌相关性研究的初步尝试,准确了解我国热带地区树种对木腐真菌分布的影响还需要开展进一步的深入研究。
总体而言,同属热带的海南尖峰岭、霸王岭和西双版纳,木腐真菌具有明显的热带区系特征。就已采集和记录的真菌种类而言,各地区物种组成存在较大差异,造成这种差异的最主要因素推测为林分中树种组成的不同。为更好地保护我国热带地区的木腐真菌资源,应有针对地围绕相关地区树种组成情况,采取不同的方案与措施。
致谢
衷心感谢北京林业大学生态与自然保护学院微生物研究所提供相关研究资料。
参考文献
Disentangling the effects of forest-stand type and dead-wood origin of the early successional stage on the diversity of wood-inhabiting fungi
DOI:10.1016/j.foreco.2016.07.011 URL [本文引用: 1]
Mycota and ecology of polypores in eastern Himalayas
Wood-decaying fungi in eastern Himalayas 3. Polypores from Laojunshan Mountains, Yunnan Province
Species diversity, taxonomy and phylogeny of Polyporaceae (Basidiomycota) in China
DOI:10.1007/s13225-019-00427-4 URL [本文引用: 1]
Fungal flora and population structure of polypores in the Great Xingan Mountains
A checklist of polypores in China
Hymenochaetaceae (Basidiomycota) in China
DOI:10.1007/s13225-010-0066-9 URL [本文引用: 1]
A revised checklist of corticioid and hydnoid fungi in China for 2010
DOI:10.1007/S10267-010-0068-1 URL [本文引用: 1]
Polypore diversity in China with an annotated checklist of Chinese polypores
DOI:10.1007/s10267-011-0134-3 URL [本文引用: 1]
Numbers of fungal species hitherto known in China
Colonization and extinction patterns of wood-decaying fungi in a boreal old-growth Picea abies forest
DOI:10.1111/j.1365-2745.2008.01411.x URL [本文引用: 1]
Independent effects of host and environment on the diversity of wood-inhabiting fungi
DOI:10.1111/jec.2018.106.issue-4 URL [本文引用: 1]
A comparison study on the tropical dipterocarp rain forests of Hainan and Xishuangbanna. Acta Botanica Boreali-occidentalia
Establishment of Xishuangbanna tropical forest dynamics plot: species compositions and spatial distribution patterns
Floral composition and ecological distribution of wood-decaying fungi in Laojunshan National Park, Southwest China
Floral composition and distribution characteristics of polypores in Northeast China
Community composition characteristics of wood-decaying fungi in temperate secondary forest ecosystems in China
Atlas of Chinese macrofungal resources
Mineralization of 14C-labelled synthetic lignin and extracellular enzyme activities of the wood-colonizing ascomycetes Xylaria hypoxylon and Xylaria polymorpha
Two wood-dwelling ascomycetes, Xylaria hypoxylon and Xylaria polymorpha, were isolated from rotting beech wood. Lignin degradation was studied following the mineralization of a synthetic [formula: see text]-labelled lignin in solid and liquid media. Approximately 9% of the synthetic lignin was mineralized by X. polymorpha during the growth on beech wood meal, and the major fraction (65.5%) was polymerized into water- and dioxan-insoluble material. Both fungi produced laccase (up to 1,200 U l-1) in an agitated complex medium based on tomato juice; peroxidase activity (<80 U l-1) was only detected for X. polymorpha in soybean meal suspension. The enzymatic attack of X. polymorpha on beech wood resulted in the formation of three fractions of water-soluble lignocellulose fragments with molecular masses of 200, 30 (major fraction) and 3 kDa, as demonstrated by high-performance size exclusion chromatography. This fragment pattern differs considerably from that of the white-rot fungus Bjerkandera adusta, which preferentially released smaller lignocellulose fragments (0.8 kDa). The finding that X. polymorpha produced large lignocellulose fragments, along with the fact that high levels of hydrolytic enzymes (esterase 630 U l-1, xylanase 120 U l-1) were detected, indicates the cleavage of bonds between the lignin and hemicellulose moieties.
Tree species diversity in abandoned swidden fields of Xishuangbanna, SW China
Effects of topographical factors on species diversity across Dacrydium pectinatum natural community in Hainan Island
Study on the distribution patterns and dynamics of main tree population of Fagaceae dominance in tropical montane rain forest in Hainan Island
Wood decay under the microscope
DOI:10.1016/j.fbr.2007.09.001 URL [本文引用: 1]
Global diversity and geography of soil fungi
Population difference of polypores in northwest and southeast of China
Forming mechanisms and ecological function of wood-decaying fungal community in forest ecosystem
Polypore diversity in South China
Comparison of polypore florae and diversity from temperate to subtropical forest zones in China
Community characteristics of a 60 ha dynamics plot in the tropical montane rain forest in Jianfengling, Hainan Island
DOI:10.17520/biods.2014157 URL
Floral composition and ecology of polypores in Hainan Province
Community structure and tree species diversity characteristics in a tropical montane rain forest in Bawangling Nature Reserve, Hainan Island
Phylogeny and taxonomy of Favolus (Basidiomycota)
Tropical flora of southern China
The Chinese tropical region has generally been recognized to be the area on the northern edge of tropical Asia, and includes southeastern Xizang (Tibet), southern Yunnan, southwestern Guangxi, southern Taiwan, and Hainan Island. Based on present floristic records and data from these tropical areas, 12,844 species of seed plants including 2,181 genera and 227 families, are recognized. The families that are distributed mainly in tropical areas but extended to the temperate zone contribute to the majority of the flora of tropical China, and genera with tropical distributions make up the most of the total flora, which indicate that the flora of tropical areas in China is marginally tropical in nature. The genera with the tropical Asian distribution contributed to the highest portion among the various distribution types, which implies tropical Asian or Indo-Malaysia affinity of the tropical flora of China. The tropical flora of China has conspicuous variations in floristic composition and geographical elements from region to region due to different geological history and ecological environments, although the floristic similarities at the family and generic levels are more than 90% and 64%, respectively but lower than 50% at the specific level, among the compared regional floras from southwestern China and southeastern China. We found that there are more similar dominant families and genera, and also higher similarities between families and genera between southeastern Tibet (Xizang) and southeastern Yunnan. The floras of southern and southeastern Yunnan have a higher portion of the tropical Asian elements compared with other tropical floras in China, and although they have the highest similarity at specific level, the dominant families and genera have conspicuous differences between them. The flora of Hainan has the highest ratio of tropical elements, of which the pan-tropical element has the highest portion. Differences in characteristics and evolution in these tropical floras could be influenced mainly by historical events occurring with uplift of Himalayas, such as the southeastward extrusion of the Indochina geoblock, clockwise rotation and southeastward movement of Lanping-Simao geoblock, divergent geological histories between southern and southeastern Yunnan, and southeastward movement of Hainan Island.
A comparative study on the tropical rain forests in Xishuangbanna and Hainan
西双版纳热带森林动态监测样地——树种组成与空间分布格局
西双版纳刀耕火种弃耕地树种多样性比较研究
在西双版纳热带地区选择了4块不同演替年龄的刀耕火种弃耕地(4年、9年、29年、大约40年),通过“空间代替时间”的办法,采用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(E)、Margalef丰富度指数(D)分别测定其树种多样性,并进行了比较研究。结果表明,群落的演替阶段不同,树种组成及其多样性也有明显差异。在重要值最大的前5个树种中,山黄麻(Trema orientalis)群落全部都为先锋树种组成;中平树(Macaranga denticulata)群落仍以先锋树种占优势,但
海南以壳斗科植物为优势的山地雨林主要乔木种群分布格局及动态研究
我国温带至亚热带多孔菌区系和多样性比较
海南尖峰岭热带山地雨林60ha动态监测样地群落结构特征
DOI:10.17520/biods.2014157
[本文引用: 2]

热带山地雨林是海南岛最占优势的植被类型, 以往对热带山地雨林的研究通常基于小尺度, 缺乏大尺度上的长期固定样地监测。作者基于海南岛尖峰岭地区60 ha (1,000 m × 600 m)植被动态监测大样地, 详细描述了该样地所属典型热带山地雨林的群落结构特征, 以期为更深入地揭示我国热带雨林地区的物种多样性及其维持机制提供背景资料。尖峰岭大样地20 m × 20 m样方水平上海拔和坡度分别在866.3-1,016.7 m和1.7°-49.3°间变化。大样地内共记录到439,676株存活的胸径≥1.0 cm的乔灌木植株, 除61个植株未确定种名外, 其他分属于62科155属290种。单位面积植株密度为0.7328株/m<sup>2</sup> (含萌条和分枝), 20 m × 20 m尺度上单位面积物种数量为32-120种, 平均80种。植物属区系成分以热带性质为主, 共有136属, 占总属数的88.3%; 另有鼠李科鼠李属(Rhamnus)为世界分布, 木兰科拟单性木兰属(Parakmeria)为中国特有分布。按物种重要值排序, 大样地内最具优势(重要值大于2.0)的6个种分别为: 大叶蒲葵(Livistona saribus)、白颜树(Gironniera subaequalis)、厚壳桂(Cryptocarya chinensis)、油丹(Alseodaphne hainanensis)、四蕊三角瓣花(Prismatomeris tetrandra)和海南韶子(Nephelium topengii)。优势科为樟科、壳斗科、茜草科和棕榈科, 重要值均超过了5.0。取样面积达到7.5 ha或取样数量达到2万株时, 计数到的物种数量达到261种, 占总物种数量的90.0%。大样地内稀有种和偶见种各占所有物种的20.7%和37.6%。样地内所有个体平均胸径5.22 cm, 植株径级分布呈明显的倒“J”形。本文为后续尖峰岭大样地的研究提供了背景资料, 尖峰岭大样地的建立也为热带地区生物多样性长期变化监测提供了一个基础平台。
海南岛霸王岭热带山地雨林群落结构及树种多样性特征的研究
以海南岛霸王岭自然保护区1 hm2老龄原始林样地的调查材料为基础,分析了热带山地雨林群落的组成、高度结构、径级结构及有关的树种多样性特征。结果表明:霸王岭热带山地雨林树种较丰富,物种多样性指数较高。树种数和树木的密度都随高度级、径级的增加而呈负指数或负幂函数递减;热带山地雨林不同高度级、不同径级和不同小样方斑块内的树种数都与树木密度呈显著的正相关关系。热带山地雨林经过自然的演替达到老龄顶极群落后,最后进入主林层的只是少部分树种的少数个体。
中国南部热带植物区系
DOI:10.17520/biods.2016055
[本文引用: 1]
中国的热带地区包括西藏东南部、云南西南到东南部、广西西南部、广东雷州半岛、台湾的南部和海南岛。依据现有植物区系的记录和资料, 中国的热带地区至少具有野生种子植物227科2,181属12,844种。中国的热带植物区系以热带和主产热带, 但分布区延伸到亚热带和温带的科为主, 在属的组成上, 也以热带分布属占优势, 并以热带亚洲分布属所占比例最大, 标志着其热带边缘性质和具有热带亚洲植物区系的特点。因不同地区地质历史及生态环境的差异, 中国热带植物区系在不同地区的组成和地理成分也有一定差异。总的来说, 我国西南部到东南部各热带地区的植物区系科和属的相似性分别在90%和64%以上, 但种的相似性一般低于50%。具体而言, 西藏东南部与云南东南部显示出具有更多的共同优势科属和更大的植物区系相似性; 云南南部和东南部热带亚洲成分比例最高, 虽然它们之间在种的相似性上最大, 但在优势的科属上差异较大; 海南植物区系热带成分总体比例最高, 其中又以泛热带分布比例最大。中国热带植物区系在不同地区间演化和发展上的差异与喜马拉雅隆升过程中发生的地质事件有关, 如印度支那板块向东南逃逸、云南发生地质板块顺时针旋转和位移、云南南部与东南部在地质历史上曾有的隔离以及海南岛向东南的位移等。
西双版纳热带雨林与海南热带雨林的比较研究



{kind=link}
{kind=link}
{kind=link}
{kind=link}