



“苇菇”又称“红柳菇”,生长在新疆等西北内陆湖畔,由于子实体个体肥厚、味道鲜美,是当地著名的野生食用菌。“苇菇”生长于芦苇丛或红柳林中,子实体埋生,一般春秋两季出菇,每年4-5月间为出菇旺季。根据“苇菇”子实体菌褶离生、孢子印棕色和具有菌环的形态特征,可以明确它属于蘑菇属Agaricus L.,隶属于担子菌门Basidiomycota、蘑菇纲Agaricomycetes、蘑菇目Agaricales、蘑菇科Agaricaceae。蘑菇属物种被广泛认为是草腐性的,而且世界性分布,能适应各种不同生态和气候类型,从靠近北极圈的寒温带到温带、热带亚热带地区;从草原、森林到干旱的荒漠地区都有分布。蘑菇属物种的形态多样,大多数具有典型的伞菌形态,为了适应不同的生态和气候类型,少数物种腹菌化,如在澳大利亚中部干旱区分布的A. colpeteorum T. Lebel、A. lamelliperditus T. Lebel & M.D. Barrett、A. wariatodes (Grgur.) T. Lebel和A. chartaceus T. Lebel(Parra 2008,2013;Kerrigan 2016;He et al. 2017)。
蘑菇属分类研究历史悠久,分类系统繁多,近年来随着分子技术的发展并在分类研究中广泛应用,蘑菇属的分类系统也不断趋向于自然和稳定。Zhao et al. (2016)在全球蘑菇属标本的多基因系统发育与形态学研究基础上将演化时间作为新增指标,建立了蘑菇属标准化分类体系,将蘑菇属划分为5个亚属以及20个组。在随后多项研究中,采用蘑菇属内演化时间在30 Mya左右分亚属、20 Mya左右分组为补充指标,又有来自亚洲及美洲的新亚属和组发表(Chen et al. 2017;He et al. 2018;Parra et al. 2018;Ortiz-Santana et al. 2021),目前蘑菇属共包括6个亚属和26个组。这些工作为蘑菇属资源的准确认识提供了必要的保障。
蘑菇属的种类丰富,第10版《真菌词典》记载全世界蘑菇属共约200个物种(Kirk et al. 2008),近年来由于其分子系统发育研究及分类系统的不断完善,大量新种被发现,蘑菇属物种数量已达500余种(He et al. 2019),目前我国已报道的有分子系统发育和形态学研究共同支持的蘑菇属物种116个(刘安琪 2020)。蘑菇属物种具有较高的食用和保健价值,我国共有47个种可食用,其中18种兼具食用与药用价值(Wu et al. 2019),随着蘑菇属资源的不断发现及研究的深入,我国蘑菇属的食药用菌种类数量仍在不断增加(Cao et al. 2021)。
本研究对新疆埋生或半埋生,俗称为“苇菇”的蘑菇属物种进行了较为广泛的采样,经过形态和分子系统学研究,发现“苇菇”并不是单一物种,而包含了蘑菇属4个物种,分别为大肥蘑菇A. bitorquis (Quél.) Sacc.、中华美味蘑菇A. sinodeliciosus Z.R. Wang & R.L. Zhao、帕达纳蘑菇A. padanus Lancon.和亚托柄蘑菇A. subperonatus (J.E. Lange) Singer,其中中华美味蘑菇A. sinodeliciosus是2015年发表来自新疆的新种。通过文献调研,这4个种中只有中华美味蘑菇仅在我国有报道,其他3个种在我国和欧洲等地均有分布。据此,依据中华美味蘑菇已知的地理分布数据和分布地的环境因子数据,以最大熵模型(MaxEnt)对其可能的分布范围进行了模拟预测,为其保护和可持续利用提供理论基础。
1 材料与方法
1.1 研究材料
91份标本为新疆维吾尔自治区采集获得,保存于中国科学院微生物研究所菌物标本馆(HMAS)。
1.2 物种鉴定
1.2.1 形态学研究
1.2.2 分子系统学研究
DNA提取、序列扩增和测序、序列处理及系统发育分析等参照曹槟等(2019)的方法。
1.3 中华美味蘑菇分布区预测和适生性分析
1.3.1 分布数据
数据来自2015-2019年间在新疆、青海的野外调查,以GPS记录采集点的坐标信息,共计20个中华美味蘑菇的分布样点。
1.3.2 气候数据
采用1950-2000年生物气候数据,包括19个生物气候变量(Hijmans et al. 2005)。所有的气候数据均下载于世界气象数据库Worldclim (
1.3.3 MaxEnt模型分析
采用MaxEnt version 3.4.1对分布数据进行分析,随机选取75%的样点作为训练集,其余作为验证数据集,对预测结果的准确度进行评估。利用ArcGIS对预测结果进行重分类,将预测图的适生值划分为4个等级(0-0.25为不适生区;0.26-0.50为低适生区;0.51-0.75为中适生区;0.76-1.0为高适生区)。利用刀切法(Jackknife)检验变量重要性,同时利用ROC曲线评价模型预测结果的精确度,ROC曲线下的面积(AUC)越大表示模型模拟效果越好(0.5-0.6,不及格;0.6-0.7,较差;0.7-0.8,一般;0.8-0.9,良好;0.9-1.0,优秀) (Phillips & Dudík 2008)。
2 结果与分析
2.1 分子系统发育研究结果
本研究共提取了91份“苇菇”标本DNA,并获得ITS序列,通过BLAST确认了“苇菇”序列均属于蘑菇属Pseudochitonia亚属的Bivelares组和Nigrobrunnescentes组。从GenBank中下载了Pseudochitonia亚属32个种的参考序列54条(其中模式标本序列12条),涵盖了Bivelares组和Nigrobrunnescentes组的已知物种。选取自测及GenBank下载的共计146条蘑菇属ITS序列,以草地蘑菇Agaricus campestris L.作为外群,采用最大似然法构建系统发育树(图1)。根据系统发育树,“苇菇”标本分布在Pseudochitonia亚属的Bivelares组和Nigrobrunnescentes组中。29份标本和大肥蘑菇A. bitorquis、32份标本和中华美味蘑菇A. sinodeliciosus以及3份标本和亚托柄蘑菇A. subperonatus聚在一起,分别获得99%、100%以及87%的自展支持率,并同属于Bivelares组获得100%支持率;27份标本和帕达纳蘑菇A. padanus聚在一起,获得98%的支持率,属于Nigrobrunnescentes组,具有56%的支持率。
图1
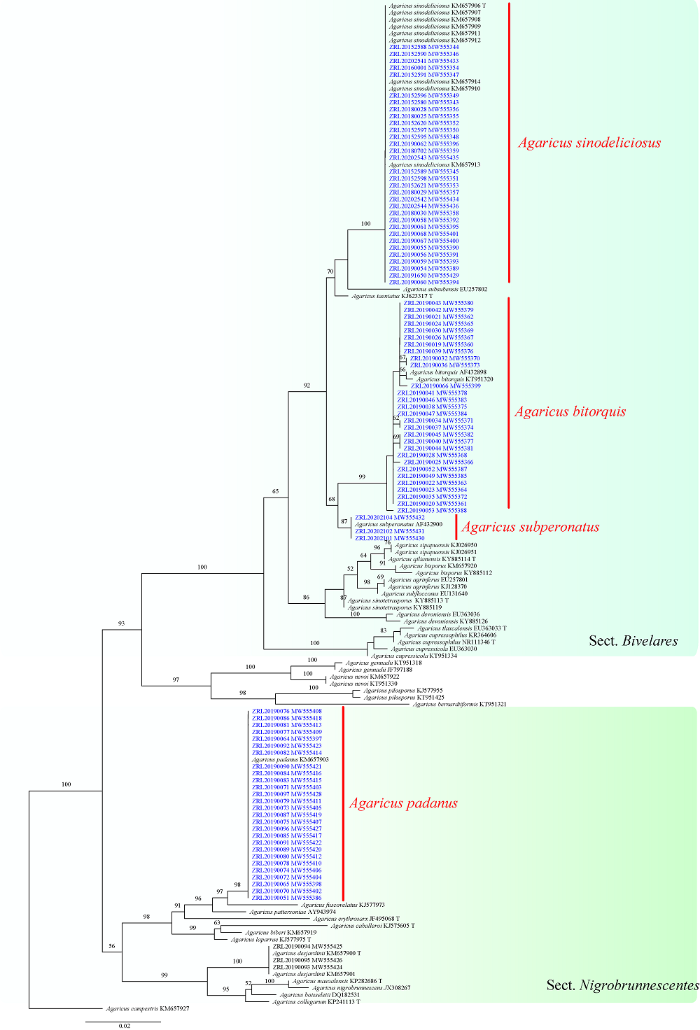
图1
基于ITS序列构建的蘑菇属Pseudochitonia亚属系统发育树(BS≥50)
Fig. 1
Phylogenetic tree of Agaricus subgenus Pseudochitonia based on ITS (BS≥50).
2.2 形态学研究结果
“苇菇”的生长环境均为气候干旱、昼夜温差大、日照时间长的盐碱沙土地,子实体埋生或半埋生(图2A)。构成“苇菇”的4个种宏观形态特征相似,如菌盖初半球形,后扁半球形至圆盘形,表面具有紧贴的棕色鳞片;菌褶离生,幼时为白色,随着成熟过程变为粉红色、红棕色至棕黑色;菌肉白色,伤后变淡红色或红棕色。在显微镜下,它们的微观特征也具有很多的相似性,如担孢子均为椭球形,光滑,深棕色;担子棍棒状,具4个担子小梗;褶缘囊体多为棒状;菌盖表皮菌丝圆柱形,偶尔分支,分隔处不缢缩或略微缢缩(图3)。通过细致的比较形态学研究,结合系统发育分析结果,发现构成“苇菇”的实际为4个种,且区分的关键形态特征有:菌环发育方式及特征、菌褶侧生囊体有无以及担孢子大小。据此,“苇菇”的形态分种检索表如下:
1. 菌环从下向上发育,往往开口向上,靴状(inferous) 2
1. 菌环从上向下发育,往往开口向下,裙状(superous) 3
2. 双菌环;菌褶无侧生囊体;担孢子小,5.5-5.9- 6.7×4.5-4.9-5 μm 大肥蘑菇A. bitorquis
2. 单菌环;菌褶有侧生囊体;担孢子大,6.4-7.2-8.3×4.9-5.7-6.4 μm 中华美味蘑菇A. sinodeliciosus
3. 菌环厚;担孢子小,5.5-6.3-7.5×4.8-5.2-6 μm 亚托柄蘑菇A. subperonatus
3. 菌环薄,易破碎;担孢子大,6.5-7.7-9.2×5- 6.1-7.2 μm 帕达纳蘑菇A. padanus
图2

图2
“苇菇”子实体宏观形态图 A:子实体埋生;B:大肥蘑菇;C:中华美味蘑菇;D:亚托柄蘑菇;E:帕达纳蘑菇. 标尺=2 cm
Fig. 2
Basidiomes. A: Buried furit bodies; B: Agaricus bitorquis; C: A. sinodeliciosus; D: A. subperonatus; E: A. padanus. Bars=2 cm.
图3
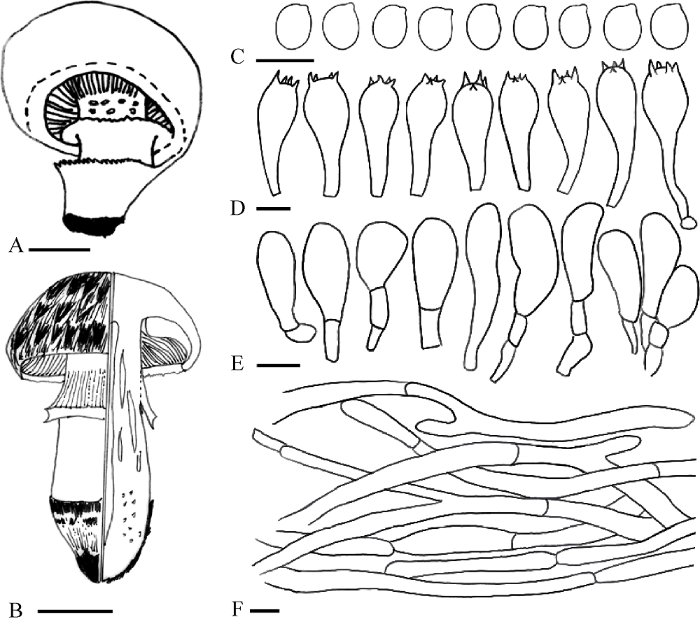
图3
“苇菇”的关键形态特征 A:大肥蘑菇(菌环从下向上发育,开口向上,靴状);B:亚托柄蘑菇(菌环从上向下发育,开口向下,裙状);C:担孢子形态;D:担子形态;E:褶缘囊体形态;F:菌盖表皮菌丝形态. 标尺:A,B=2cm,C-F=10μm. A和B参考Parra (2008)
Fig. 3
Key characteristics of ‘reed mushrooms’. A: Agaricus bitorquis (Annulus inferous); B: A. subperonatus (Annulus superous); C: Basidiospores; D: Basidia; E: Cheilocystidia; F: Pileipellis hyphae. Bars: A, B=2 cm, C-F=10 μm. A and B were redrawn by referring to Parra (2008).
Agaricus subgenus Pseudochitonia section Bivelares
Agaricus bitorquis (Quél.) Sacc., Syll. Fung. 5: 998 (1887) Fig. 2B
大肥蘑菇 图2B
子实体埋生、半埋生或表生。菌盖直径40-150 mm,初半球形,后扁半球形至圆盘形,顶部平或中心略下凹,表面光滑,白色,受伤后变淡黄色。菌褶离生,密集,宽达5 mm,幼时为白色、粉红色,成熟后为深棕色。菌柄长40-120 mm,直径12-30 mm,等宽或基部略微膨大,表面光滑,白色,受伤后变红棕色。菌环从下往上发育,开口向上,靴状,往往形成双菌环,膜质,通常生于菌柄下部。菌肉白色,伤后变淡红色或红棕色。气味蘑菇味。
担孢子5.5-5.9-6.7(-7)×(4-)4.5-4.9-5(-5.5) μm,Q=1.1-1.21-1.35(-1.4),近球形至椭球形,光滑,深棕色。担子20-26×(5-)7-10 μm,棍棒状,具4个担子小梗,小梗长达3 μm。褶缘囊体12-30× 6-13 μm,多为棒状。菌盖表皮菌丝宽3-9 μm,分隔处略有缢缩,端部渐细。
KOH和Schäffer反应:阴性(不变色)。
分布:世界广布。
研究标本:新疆巴音郭楞蒙古自治州博湖县,2019年6月8日,曹槟,HMAS 281628 (ZRL20190019),HMAS 291221-HMAS 291227 (ZRL20190020-ZRL20190026),HMAS 281629 (ZRL20190028),HMAS 291229 (ZRL20190030),HMAS 291230 (ZRL20190032),HMAS 291231- HMAS 291244 (ZRL20190034-ZRL20190047)。新疆巴音郭楞蒙古自治州和硕县,2019年6月12日,曹槟,HMAS 291245 (ZRL20190049),HMAS 291247 (ZRL20190052),HMAS 291248 (ZRL20190053)。新疆博尔塔拉蒙古自治州博乐市新疆生产建设兵团农五师九十团,2019年6月14日,曹槟,HMAS 255776 (ZRL20190066)。
Agaricus sinodeliciosus Z.R. Wang & R.L. Zhao, Phytotaxa 202(3): 192 (2015) Fig. 2C
中华美味蘑菇 图2C
子实体埋生或半埋生。菌盖直径50-120 mm,初为垫状或扁球形,之后展开菌盖中央略微内陷,边缘内卷,表面覆盖紧贴的细小棕色或淡黄色的鳞片,底色为污白色,受伤后变红棕色。菌褶离生,密集,较窄,红棕色至棕色,完全成熟后为深棕色,且有多种小菌褶。菌柄长20-100 mm,直径30-80 mm,初为宽梭形并具有钝圆的基部,成熟后为圆柱形并具有逐渐尖锐的基部。菌环为较厚的膜质,自下向上发育,开口朝上,靴状,下表面棕黄色。菌肉厚实,白色,伤后变红棕色。气味蘑菇味。
担孢子6.4-7.2-8.3(-9)×4.9-5.7-6.4 μm,Q=1.2-1.27-1.5,宽椭球形至椭球形,光滑,棕色。担子数量少,13-17×4-7 μm,棒状,具4个担子小梗,小梗长达2 μm。褶缘囊体20-41× 6-10 μm,幼时长椭圆形,逐渐发育为棒状或宽棒状,通常具有一个细长的柄部,透明或内部填充棕黄色色素。具侧生囊体,形态与褶缘囊体相似。菌盖表皮菌丝宽4-16 μm,圆柱状,有分支,弯曲,越宽的菌丝其分隔处越窄,内部含有淡棕色的色素。
KOH和Schäffer反应:阴性(不变色)。
分布:中国新疆、青海等西北地区。
研究标本:新疆博尔塔拉蒙古自治州精河县艾比湖,2012年5月10日,王卓仁,HMAS 255793 (Isotype)。新疆巴音郭楞蒙古自治州和硕县,2019年6月12日,曹槟,HMAS 255773- HMAS 255775 (ZRL20190054-ZRL20190056),HMAS 281630 (ZRL20190058),HMAS 281425 (ZRL20190059),HMAS 281631 (ZRL20190060),HMAS 291249 (ZRL20190061),HMAS 291250 (ZRL20190062),HMAS 291253 (ZRL20190067),HMAS 281632 (ZRL20190068)。
Agaricus subperonatus (J.E. Lange) Singer, Lilloa 22: 432 (1951) Fig. 2D
亚托柄蘑菇 图2D
子实体埋生、半埋生或表生。菌盖直径50-150 mm,初半球形,后平展至圆盘形,表面具有紧贴的三角形纤维状鳞片,鳞片呈深棕色或红棕色,菌盖边缘通常超出菌褶达5 mm。菌褶离生,密集,宽达7 mm,幼时为白色成熟后为深棕色。菌柄长35-150 mm,直径15-30 mm,棒状或圆柱形,基部渐细外菌幕常残留,菌环以上部分光滑,白色,菌环以下具纤维状覆盖物。菌环为较厚的膜质,自上向下发育,开口朝下,裙状,上表面具浅白色条纹,下表面具纤维状覆盖物。菌肉白色,伤后变淡红色。气味蘑菇味。
担孢子(5.2-)5.5-6.3-7.5(-8.2)×(4.5-)4.8-5.2- 6(-7) μm,Q=1.1-1.21-1.35,近球形至椭球形,光滑,深棕色。担子25-36×8-14 μm,棍棒状,担孢子小梗长达4 μm。褶缘囊体21-53×7-13 μm,多为棒状或圆柱形,极少梭形。菌盖表皮菌丝宽4-11 μm,分隔处几乎不缢缩。
KOH和Schäffer反应:阴性(不变色)。
分布:中国西北地区;欧洲广布;北美五大湖周边。
研究标本:新疆昌吉回族自治州玛纳斯县兰州湾镇,2020年9月,林汝楷,HMAS 291530 (ZRL20202101),HMAS 291531 (ZRL20202102),HMAS 291532 (ZRL20202104)。
Agaricus subgenus Pseudochitonia section Nigrobrunnescentes
Agaricus padanus Lancon., Riv. Micol. 45(1): 30 (2002) Fig. 2E
帕达纳蘑菇 图2E
子实体埋生、半埋生或表生。菌盖直径40-80 mm,初半球形,后圆盘形,中心通常内陷,表面覆盖淡棕色至黄棕色鳞片,底色为灰白色,菌盖触碰后变红棕色。菌褶离生,密集,棕色至深棕色,边缘颜色浅且完整。菌柄长40- 80 mm,直径20-38 mm,表面光滑,白色至灰白色。菌环自上向下发育,开口朝下,裙状,单层,膜质,上表面条纹状。菌肉伤后初为淡灰白色,逐渐变淡红色或紫罗兰色。气味蘑菇味。
担孢子6.5-7.7-9.2×5-6.1-7.2 μm,Q=1.07- 1.27-1.55,近球形至椭球形,光滑,棕色。担子18-30×6-12 μm,棒状顶部圆或平截,透明具有2或4个担子小梗,小梗长达4 μm。褶缘囊体15-30×6-16 μm,棒状或端部膨大或不规则的棒状,基部通常具有1-2个分隔,透明或具有淡棕色的色素。菌盖表皮菌丝宽3-7 μm,圆柱形,偶尔分支,分隔处不缢缩或略微缢缩。
KOH和Schäffer反应:阴性(不变色)。
分布:中国西北地区;意大利。
研究标本:新疆巴音郭楞蒙古自治州和硕县,2019年6月12日,曹槟,HMAS 291246 (ZRL20190051)。新疆博尔塔拉蒙古自治州博乐市新疆生产建设兵团农五师九十团,2019年6月14日,曹槟,HMAS 291251 (ZRL20190064),HMAS 291252 (ZRL20190065)。新疆博尔塔拉蒙古自治州博乐市新疆生产建设兵团农五师九十一团,2019年6月14日,曹槟,HMAS 281633 (ZRL20190070),HMAS 291254-HMAS 291257 (ZRL20190071-ZRL20190074),HMAS 281634 (ZRL20190075),HMAS 291258-HMAS 291260 (ZRL20190076-ZRL20190078),HMAS 281635 (ZRL20190079),HMAS 291261 (ZRL20190080),HMAS 291262 (ZRL20190081),HMAS 255777 (ZRL20190082),HMAS 255778 (ZRL20190083),HMAS 291263-HMAS 291266 (ZRL20190084- ZRL20190087)。新疆博尔塔拉蒙古自治州博乐市新疆生产建设兵团农五师八十二团蘑菇滩,2019年6月15日,曹槟,HMAS 291267-HMAS 291270 (ZRL20190089-ZRL20190092),291273 (ZRL20190096),HMAS 291274 (ZRL20190097)。
2.3 中华美味蘑菇的生态分布区预测
2.3.1 模型评估
计算结果显示,平均训练和测试AUC值分别为0.998和0.999,说明MaxEnt模型模拟效果非常好。
2.3.2 潜在分布范围
中华美味蘑菇适生区较为狭窄,高适生区主要位于我国西北部及中亚地区(哈萨克斯坦及蒙古国),在我国主要分布于新疆天山山脉南北两侧区域、西藏西南部、青海海西州、甘肃西北部以及内蒙古西部地区(图4)。
图4
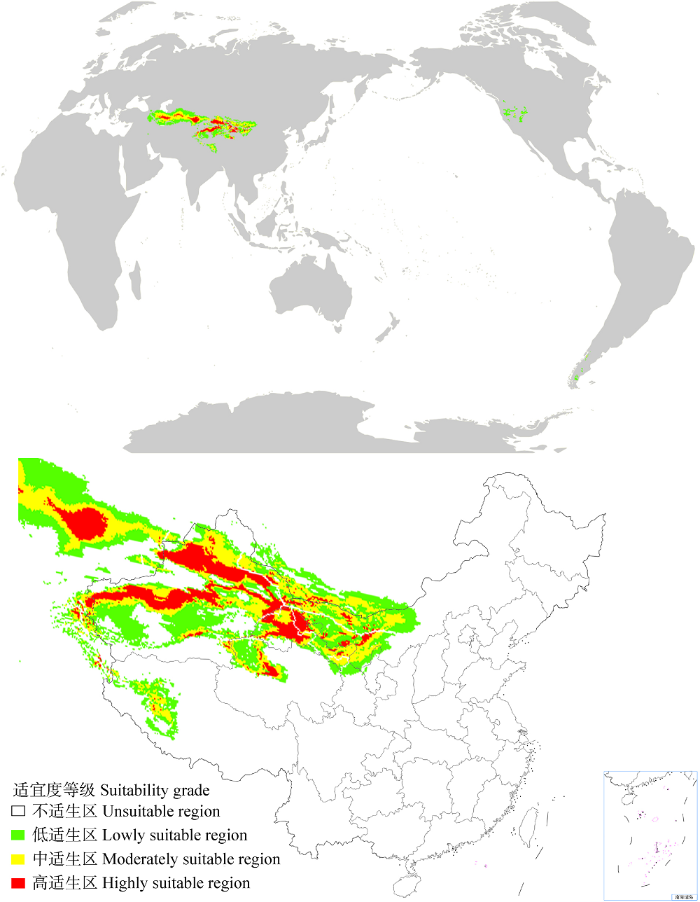
图4
基于MaxEnt模型预测中华美味蘑菇的适生区
审图号:GS (2021) 6081号
Fig. 4
Potential distribution regions of Agaricus sinodeliciosus.
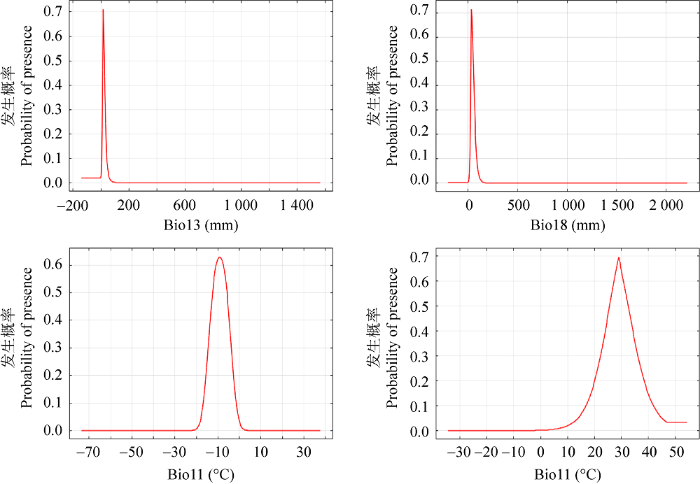
2.3.3 影响中华美味蘑菇分布的主要环境因子
表1 物种分布模型涉及的环境变量及其对模型的贡献率
Table 1
| 编号 Code | 环境变量 Environmental variable | 贡献率 Contribution (%) |
|---|---|---|
| Bio1 | 年均温Annual mean temperature | 1.0 |
| Bio2 | 昼夜温差月均值Mean diurnal range | 0.5 |
| Bio3 | 等温性Isothermality | 3.3 |
| Bio4 | 温度季节性变化的标准差Temperature seasonality | 0 |
| Bio5 | 最暖月最高温Max. temperature of warmest month | 15.4 |
| Bio6 | 最冷月最低温Min. temperature of coldest month | 1.0 |
| Bio7 | 年均温变化范围Temperature annual range | 0.2 |
| Bio8 | 最湿季度平均温度Mean temperature of wettest quarter | 0 |
| Bio9 | 最干季度平均温度Mean temperature of driest quarter | 0 |
| Bio10 | 最暖季度平均温度Mean temperature of warmest quarter | 0 |
| Bio11 | 最冷季度平均温度Mean temperature of coldest quarter | 16.5 |
| Bio12 | 年均降水量Annual precipitation | 0 |
| Bio13 | 最湿月降水量Precipitation of wettest month | 40.0 |
| Bio14 | 最干月降水量Precipitation of driest month | 2.2 |
| Bio15 | 降水量变异系数Precipitation seasonality | 0.6 |
| Bio16 | 最湿季度降水量Precipitation of wettest quarter | 0 |
| Bio17 | 最干季度降水量Precipitation of driest quarter | 1.1 |
| Bio18 | 最暖季度降水量Precipitation of warmest quarter | 16.9 |
| Bio19 | 最冷季度降水量Precipitation of coldest quarter | 1.3 |
图5
3 讨论
3.1 “苇菇”的物种多样性
通过较为广泛的标本采集和相关研究表明,生长于新疆芦苇及红柳林中,子实体埋生、半埋生的野生食用菌“苇菇”虽然形态特征较为相似,但并不是单一物种,而是分属于蘑菇属Pseudochitonia亚属,2个不同组的物种,分别为Bivelares组的大肥蘑菇A. bitorquis、中华美味蘑菇A. sinodeliciosus和亚托柄蘑菇A. subperonatus;及Nigrobrunnescentes组的帕达纳蘑菇A. padanus。这4个物种为了适应气候干旱、昼夜温差大、日照时间长的盐碱沙土地(王秀云和孔爱琴 1987;张连成和王勇辉 2014),子实体表现出埋生或半埋生的特性。其中只有中华美味蘑菇A. sinodeliciosus现仅在我国有报道(Wang et al. 2015;徐鸿雁等 2018;胡日瓦 2019;徐珍等 2019);而大肥蘑菇A. bitorquis是世界广布种,能适应各种不同生态和气候类型,也有埋生的报道(Parra 2008;Kerrigan 2016);帕达纳蘑菇A. padanus除分布在我国外(胡日瓦和图力古尔 2019),仅在意大利有报道,主要生长在运河两边的草地上(Parra 2008);亚托柄蘑菇A. subperonatus欧洲广布(Parra 2008),在北美主要分布于五大湖周边,也有埋生或半埋生的情况(Kerrigan 2016),在我国已有文献记载(曾辉等 1995),在本研究中首次获得分子系统发育支持,并保藏了凭证标本。本文的研究结果表明过去对我国西部地区大型真菌种类研究薄弱(戴玉成等 2021;王科等 2021),西部地区大型真菌的多样性还有待进一步研究。
中华美味蘑菇发表于2015年,但之前通过形态学研究,发现该物种形态特征与原产地为欧洲的圆孢蘑菇A. gennadii (Chatin & Boud.) P.D. Orton相似,所以中华美味蘑菇曾被命名为圆孢蘑菇(卯晓岚 2000)。随着分子生物学的发展,以DNA序列为基础的分子系统发育研究表明来自新疆的“圆孢蘑菇”和欧洲的圆孢蘑菇分属于蘑菇属中亲缘关系较远的不同组内:新疆标本属于蘑菇属的Bivelares组,而来自欧洲的圆孢蘑菇则属于Chitonioides组(Wang et al. 2015)。在形态学上这两个种也存在明显区别:新疆的“圆孢蘑菇”具有中到大型的子实体,多为埋生或半埋生,菌肉受伤后有明显的红棕色变色,最为特殊的是菌褶除了拥有褶缘囊体外,还有侧生囊体(Wang et al. 2015);而圆孢蘑菇并不具有埋生或半埋生的生长特性,菌肉受伤后不变色,也没有侧生囊体(Parra 2008)。因此来自新疆的“圆孢蘑菇”被正式描述并命名为中华美味蘑菇A. sinodeliciosus,模式标本保存于中国科学院微生物研究所菌物标本馆,模式标本的ITS序列提交GenBank (No. KM657906) (Wang et al. 2015)。
此外在GenBank中的ITS序列No. AM930983和中华美味蘑菇相似度非常高,而这个菌株被给予的名字为原产地来自哈萨克斯坦的巴尔喀什蘑菇A. balchaschensis Samgina & G.A. Nam (Hildén et al. 2013)。通过查阅发表该序列的论文发现,该论文中没有任何形态描述和相关分类信息,无法确定该菌株鉴定为巴尔喀什蘑菇的分类依据。然而根据1989年发表巴尔喀什蘑菇的原始论文(这也是关于巴尔喀什蘑菇唯一的分类学论文),中华美味蘑菇和巴尔喀什蘑菇有明显的形态差异,如巴尔喀什蘑菇没有褶缘囊体和侧生囊体(Samgina & Nam 1989),而中华美味蘑菇拥有大量的褶缘囊体和侧生囊体等。所以我们推测GenBank中序列AM930983很有可能应该为中华美味蘑菇,但被错误鉴定为巴尔喀什蘑菇。
3.2 中华美味蘑菇适生区预测表明其为狭域种
本研究调查发现,近年来因过度采挖和生态环境改变等因素影响,中华美味蘑菇野生资源已日趋减少。如博斯腾湖曾经为中华美味蘑菇的主产区,但自从2016年以来,湖水持续上涨,造成周边芦苇、红柳面积不断缩小,从而使其所需生境面积日趋减少,博斯腾湖周边中华美味蘑菇种群已濒临灭绝。中华美味蘑菇适生区的预测结果不仅对于全面掌握中华美味蘑菇资源的分布状况具有一定参考价值,同时也能为中华美味蘑菇种质资源保护及人工抚育区域规划提供必要的理论参考。
3.3 中华美味蘑菇资源保护、挖掘与可持续利用
在就地保护方面,随着政府部门对生态环境、生物多样性的重视,2000年6月中华美味蘑菇栖息地之一的艾比湖湿地成为自治区级自然保护区,2007年4月又晋升为国家级自然保护区,中华美味蘑菇的栖息地受到了一定的保护。虽然保护区未将中华美味蘑菇作为主要保护对象,但在保护生态环境的同时,也在一定程度上保护了中华美味蘑菇。在迁地保护方面,对中华美味蘑菇开展人工驯化栽培技术的研究,是近年来食用菌领域的一个关注重点,并有人工驯化出菇的报道,如李传华等(2018)发现中华美味蘑菇不需覆土即可出菇。虽然中华美味蘑菇已实现人工实验室驯化,但大面积栽培的技术尚不成熟(王俊燕 2002;苟小清等 2014;徐鸿雁等 2018)。中华美味蘑菇作为狭域种,野生资源有限,因此还需要通过各项保护措施加大对其保护力度。
大多数食用菌都适宜在偏酸的环境中生长,适合菌丝生长的pH在3-8之间,以5-6.5为宜,pH大于7时生长受阻,大于9时几乎完全停止生长,不同的食用菌其最适pH稍有不同,双孢蘑菇喜中性环境,灰树花最耐酸,其他大都喜偏酸性环境(罗信昌和陈士瑜 2010),然而本文中所描述的蘑菇属物种在我国西北生态脆弱地区经历了长期的演化,是能适应特殊盐碱环境的大型真菌代表(凌志琳 2019),因此深入挖掘中华美味蘑菇耐盐碱遗传资源具有重要意义。前期通过CAPS分子标记、交配实验和出菇实验的验证,已证明中华美味蘑菇的生活史为异宗配合为主,存在部分假同宗配合,其假同宗配合遵循非随机模型(Ling et al. 2019)。这些研究对该菇的人工栽培条件优化、新品种培育、开发等方面提供了必要的理论基础。
致谢:
感谢新疆生产建设兵团农业建设第五师农业科学研究所付文进研究员和巴音郭楞州科学技术局赵春生工程师在野外调查中给予的指导和帮助。感谢青海大学农林科学院青藏高原野生植物资源研究所徐鸿雁研究员提供标本。
参考文献
A revision of Agaricus section Arvenses with nine new species from China
Species diversity and geographic components of Russula from the Greater and Lesser Khinggan Mountains
Study in Agaricus subgenus Minores and allied clades reveals a new American subgenus and contrasting phylogenetic patterns in Europe and Greater Mekong Subregion
Diversity and systematics of the important macrofungi in Chinese forests
Domestication and cultivation of Agaricus gennadii
Tropic origins, a dispersal model for saprotrophic mushrooms in Agaricus section Minores with descriptions of sixteen new species
A new section and species of Agaricus subgenus Pseudochitonia from Thailand
Notes, outline and divergence times of Basidiomycota
Very high resolution interpolated climate surfaces for global land areas
Agaricus bisporus and related Agaricus species on lignocellulose production of manganese peroxidase and multicopper oxidases
Investigation and domestication on Agaricus species in Inner Mongolia and Northeast China
Biological characteristics and domestication cultivation of wild edible fungus Agaricus padanus
Agaricus of North America
Dictionary of the fungi. 10th ed
Agaricus sinodeliciosus: one wild species of Agaricus cultivated successfully without casing
Comparative genomic and transcriptomic analysis of Agaricus sinodeliciosus
Offspring analysis using two CAPS markers reveals amphithallism in the edible mushroom Agaricus sinodeliciosus
Phylogenetic study of Agaricus in main forests of China
Collection of mushroom industry in China
The macrofungi in China
The genus Agaricus in the Caribbean II. Refined phylogeny of Agaricus subg. Spissicaules with description of two new sections and eight new species
Agaricus L. Allopsalliota Nauta & Bas. Fungi Europaei 1
Agaricus L. Allopsalliota Nauta & Bas. Fungi Europaei 1A
The genus Agaricus in the Caribbean. Nine new taxa mostly based on collections from the Dominican Republic
Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation
Novyĭ vid iz roda Agaricus L.: Fr
Experiment on domestication and cultivation of Agaricus gennadii
Overview of China’s nomenclature novelties of fungi in the new century (2000-2020)
Investigation on wild mushroom from Bosten Lake
Edible species of Agaricus (Agaricaceae) from Xinjiang Province (western China)
Resource diversity of Chinese macrofungi: edible, medicinal and poisonous species
Identification, biological characteristics and domestication of Agaricu sinodeliciosus
Domestication and cultivation of Agaricus sinodeliciosus using wheat straw substrate
Common species of Agaricus and their applications
Chemical properties of reed growing soil in Ebinur wetlands
Towards standardizing taxonomic ranks using divergence times - a case study for reconstruction of the Agaricus taxonomic system
大小兴安岭地区红菇属物种多样性及其地理成分
DOI:10.17520/biods.2019040
[本文引用: 1]

红菇属(Russula)真菌广泛分布于世界各地并与多种植物根系共生形成外生菌根, 对于森林生态系统的多样性维持与群落构建具有重要的作用。我国东北大小兴安岭地区是欧亚和美洲北温带地区生物群落交流的关键地区之一, 但该地区有关外生菌根真菌的区系研究较少。本研究对采自大小兴安岭的96份红菇标本进行了形态学鉴定和ITS序列分析, 通过结合世界范围内红菇属代表性物种(146个种)的ITS序列构建了红菇属系统发育图谱。结果表明, 大小兴安岭地区红菇属有46个分类单元, 其中包括37个种和9个未定种。对大小兴安岭地区37种红菇的地理成分分析表明, 北温带分布成分有13种、温带-热带共有成分8种、欧亚成分9种、世界广布成分7种。本研究表明大小兴安岭地区的红菇具有丰富的物种多样性, 而且具有明显的北温带区系特征, 同时也具有一定的区域特殊性。
中国森林大型真菌重要类群多样性和系统学研究
新世纪中国菌物新名称发表概况(2000-2020)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}