



生物钟(circadian clock)存在于各种真核生物中,调控多种生理活动和分子活性。生物钟是基于转录-翻译负反馈环的自主振荡器,在时间上协调从基因转录到代谢的一系列过程,已经发现存在于动物、植物、真菌和部分原核生物中(Dunlap 1999;Bell-Pedersen et al. 2005)。昼夜节律振荡器(circadian oscillator)是生物钟的核心组成,生物钟由一个或多个昼夜节律振荡器组成,它对生物钟起调节控制作用,对昼夜节律的变化有影响,具有使生物体与外界环境保持同步的功能(Li 1983)。通常生物钟生成一个24 h的持续性程序,适应环境因子发生的变化,从分子水平上反应对环境中的光和温度的影响,并介导与环境保持同步(Ripperger & Schibler 2001)。
在负反馈环中,钟蛋白FRQ和FRH的复合物FFC作为环中的负调控因子,而蓝光受体蛋白WC-1和WC-2通过PAS (PER-ARNT-SIM)域结合成复合物WCC成为环中的正调控因子。行使功能的时候,在持续的黑暗中,WCC结合到钟基因frq启动子的2个顺式元件上,以激活frq的转录(Crosthwaite et al. 1997;Talora et al. 1999;Cheng et al. 2001b;Cheng et al. 2002;Cheng et al. 2003b;Froehlich et al. 2003)。另一方面,FFC通过抑制WCC的活性来抑制frq转录。在FRQ进一步磷酸化和降解以后,WCC的再激活导致了frq的再激活和新周期的开始,更具体地说,随着FRQ的逐渐磷酸化,其对WCC的负调控逐渐减弱,最终随着新一轮FRQ转录周期的启动而降解(Larrondo et al. 2015)。因此,这个负反馈环产生内源性的由frq-mRNA和FRQ蛋白调节节律的近24 h昼夜振荡。除了在昼夜节律负反馈环中发挥重要作用外,WC-1也是负责昼夜节律传导和所有其他已知光反应的蓝光受体,表明了光输入和昼夜节律振荡器之间的联系(Froehlich et al. 2002;He et al. 2002;Cheng et al. 2003a;He & Liu 2005)。除了负反馈环中正调控因子和负调控因子之间的相互协调作用以外,研究也发现同二聚体FRQ通过与WCC复合物的物理相互作用来抑制自己的转录(Aronson et al. 1994;Cheng et al. 2001a;Denault et al. 2001;Froehlich et al. 2003)。
在本研究中,所用材料蛹虫草Cordyceps militaris (L.) Fr.与粗糙脉孢菌同为子囊菌,相关研究结果可以与粗糙脉孢菌的结果进行对比。蛋白酶体抑制剂MG132是一种可逆的醛基肽类特异性蛋白酶体抑制剂,能够选择性地阻断泛素-蛋白酶途径。前期研究用MG132处理蛹虫草进行了形态观察,表明其对蛹虫草的形态影响明显,直观地确定了该试剂可以用于蛹虫草中蛋白质泛素化方面的研究(丰磊等 2021)。本研究对蛹虫草中的钟基因Cmfrq、蓝光受体基因Cmwc-1和Cmwc-2进行蓝光和MG132处理后的转录水平分析。
1 材料与方法
1.1 菌种和主要试剂
蛹虫草菌种由实验室保存。蛋白酶体抑制剂MG132购于大连美仑生物技术有限公司,TRIzol购于生工生物工程(上海)股份有限公司;Fungal RNA Kit、Recombinant DNaseⅠ、第一链cDNA合成试剂盒、PrimeScript® RT reagent Kit With gDNA Eraser和SYBR® Premix Ex Taq Ⅱ均购于宝生物工程(大连)有限公司。
1.2 不同条件下蛹虫草菌丝体的培养
1.2.1 含有不同浓度MG132的PDA培养基上菌丝体的培养
将菌种接种在含有0、2、4、6、8、10、40、60和80 μmol/L MG132的PDA平板培养基上,25 ℃恒温培养箱培养,前4 d黑暗培养,第5天打开蓝光照射,培养第7天全部取样放入-60 ℃冰箱冻存。
1.2.2 黑暗和10 μmol/L MG132处理条件下菌丝体培养
将菌种接种于含有10 μmol/L MG132的PDA平板培养基上,25 ℃恒温培养箱培养;由于菌丝体培养的前4 d量比较少,因此从培养的第5天开始取样(下同),每天在同一时间取样,连续取样10 d,全部样品放入-60 ℃冰箱冻存,留待以后共同提取RNA。以不加MG132的PDA培养基上相应培养的蛹虫草菌丝体作为对照CK。
1.2.3 黑暗培养过程中2 h蓝光间断且10 μmol/L MG132处理条件下菌丝体的培养
将菌种接种在含有10 μmol/L MG132的PDA平板培养基上,25 ℃恒温培养箱培养;前4 d对菌丝体进行黑暗培养,第5天打开蓝光照射,照射2 h后关闭蓝光,部分样品直接取样,其他样品重新置于黑暗中培养,每天同一时间取样,连续取样10 d,全部样品放入-60 ℃冰箱冻存,留待以后共同提取RNA。以不加MG132的PDA培养基上相应培养的蛹虫草菌丝体作为对照CK。
1.2.4 蓝光照射和10 μmol/L MG132处理条件下菌丝体的培养
将菌种接种在含有10 μmol/L MG132的PDA平板培养基上,25 ℃恒温培养箱培养;前4 d对菌丝体进行黑暗培养,第5天开始蓝光照射并进行持续培养,从第5天开始取样(取样当天蓝光照射2 h),每天在同一时间取样,连续取样10 d,全部样品放入-60 ℃冰箱冻存,留待以后共同提取RNA。以不加MG132的PDA培养基上相应培养的蛹虫草菌丝体作为对照CK。
1.2.5 蓝光和黑暗交替且10 μmol/L MG132处理条件下菌丝体的培养
将菌种接种在含有10 μmol/L MG132的PDA平板培养基上,25 ℃恒温培养箱中培养;第6天开始蓝光照射,采用蓝光12 h-黑暗12 h-蓝光12 h-黑暗12 h交替培养48 h,期间每隔2 h取一次样,共取样24组。以不加MG132的PDA培养基上相应培养的蛹虫草菌丝体作为对照CK。
1.3 蛹虫草菌丝体和子实体总RNA的提取与cDNA 第一链的合成
-60 ℃保存样品取出后,取0.05-0.1 g样品在-60 ℃预冷的研砵中充分研磨,迅速加入1 mL TRIzol,氯仿去除蛋白质等杂质,异丙醇沉淀总RNA和75%酒精洗涤,最后获得蛹虫草总RNA。DNA的去除和荧光定量cDNA第一链的合成采用PrimeScript® RT reagent Kit With gDNA Eraser试剂盒,操作过程参照说明书。
1.4 引物设计
按照实时荧光定量PCR引物的要求,根据Cmfrq (GenBank:KF971860.1)、Cmwc-1 (GenBank:JX845417.1)和Cmwc-2 (GenBank:JX852619.1)基因序列设计引物。其中Cmfrq实时荧光定量PCR引物为FRQF (5ʹ-TGGAGCAATCGGACGACTAT C-3ʹ)和FRQR (5ʹ-ACCGCCACCTCATCACCCA C-3ʹ);Cmwc-1实时荧光定量PCR引物为WC-1F (5ʹ-TTGGCGGCAGATCAAGGAT-3ʹ)和WC-1R (5ʹ-TGCTCTGAAGGCTGAAACCC-3ʹ);Cmwc-2实时荧光定量PCR引物为WC-2F (5ʹ-GCCCTCCACC ACTACCG-3ʹ)和WC-2R (5ʹ-CGGAGTCGTCAAA GCCA-3ʹ)。以3-磷酸-甘油醛脱氢酶(glyceraldehyde- 3-phosphate dehydrogenase,GAPDH) (GenBank:FJ374269.1)为内标基因,其引物为G1 (5ʹ-GCCG AGGAAACAACAGAA-3ʹ)和G2 (5ʹ-GCAGTCGT GGCAAGGAT-3ʹ)。
1.5 Real-time PCR分析蛹虫草Cmfrq、Cmwc-1和Cmwc-2的相对转录水平
采用两步法的PCR扩增程序:95 ℃预变性20 s,95 ℃ 5 s和60 ℃ 34 s进行循环,40个循环。Real-time PCR 在Step One PCR System (杭州博日生物科技有限公司)上完成,用试剂盒SYBR® Premix Ex Taq Ⅱ进行荧光标记,按说明书进行。每个反应设置3个重复,取平均值,用ΔCT法处理数据:目标基因/管家基因=2-ΔCT,ΔCT=CT目标基因- CT管家基因,计算目的基因的相对转录水平。
2 结果与分析
2.1 蛹虫草总RNA的提取
采用TRIzol试剂对蛹虫草菌丝体进行RNA提取,提取的RNA经琼脂糖凝胶电泳后,可以清晰地看到28S和18S条带,表明所提取的RNA质量较好,无明显降解,可用于实时荧光定量PCR分析(图1)。
图1

图1
蛹虫草菌丝体总RNA的电泳分析
Fig. 1
Total RNA electrophoresis analysis of Cordyceps militaris mycelium.
2.2 不同浓度MG132和蓝光处理对Cmfrq、Cmwc-1和Cmwc-2转录水平的影响和MG132使用浓度的确定
蛹虫草菌丝体在25 ℃经过4 d黑暗培养后蓝光培养3 d,进行实时荧光PCR分析。结果表明,MG132浓度为实验的各浓度时,Cmfrq、Cmwc-1和Cmwc-2均有相应的表达(图2A-2C)。但是,Cmfrq、Cmwc-1和Cmwc-2的相对表达量与MG132浓度并无一致性。综合来看,确定后续研究中MG132处理菌丝体的浓度为10 μmol/L。
图2
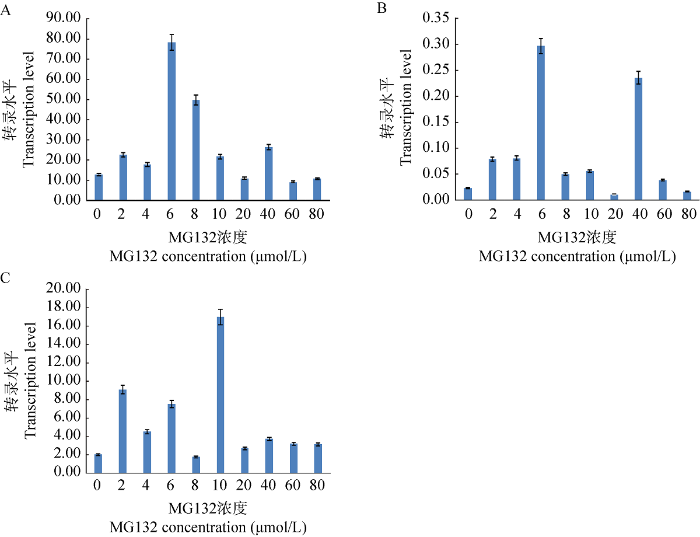
图2
不同浓度MG132和蓝光处理对Cmfrq (A)、Cmwc-1 (B)和Cmwc-2 (C)基因转录水平的影响
Fig. 2
Effects of different concentration of MG132 treatment on Cmfrq (A), Cmwc-1 (B) and Cmwc-2 (C) transcription levels under blue light irradiation.
2.3 黑暗条件下10 μmol/L MG132对蛹虫草Cmfrq、Cmwc-1和Cmwc-2转录水平的影响
蛹虫草接种在含有10 μmol/L MG132的PDA培养基上,25 ℃黑暗条件下培养至第5天开始取样,每天同一时间取样,连续取样10 d,实时荧光PCR对Cmfrq、Cmwc-1和Cmwc-2这3个基因进行转录水平分析。比较分析表明,Cmfrq基因受MG132的影响,仅在培养的第9天有一个比未加MG132的对照组高出许多的转录水平(图3A);Cmwc-1基因受MG132的影响,大多数情况下都比对照组转录水平低,但无论是加MG132的实验组还是未加MG132的对照组,转录水平均非常低,实验组和对照组的转录水平差异不明显(图3B);在取样的前2 d即培养的第5天和第6天,对照组和MG132处理组Cmwc-2基因转录均出现差异和波动,培养的第7天以后,对照组和MG132处理组的转录水平差异不大,且均处于一个较低的转录水平(图3C),说明MG132对Cmwc-2转录水平的影响有限。
图3
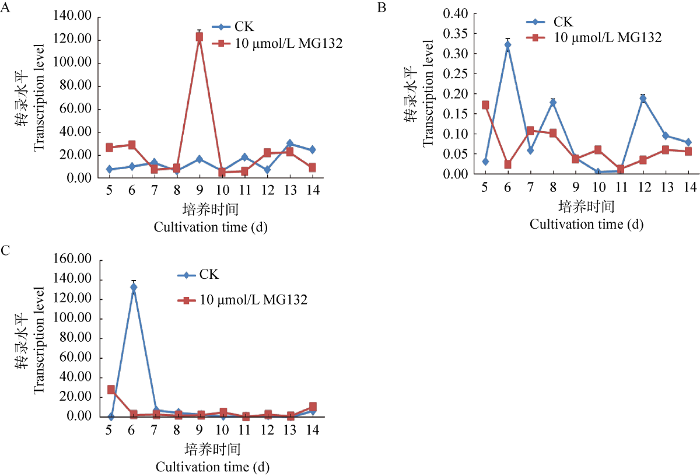
图3
黑暗条件下10 μmol/L MG132对Cmfrq (A)、Cmwc-1 (B)和Cmwc-2 (C)转录水平的影响
Fig. 3
Effects of 10 μmol/L MG132 treatment on Cmfrq (A), Cmwc-1 (B) and Cmwc-2 (C) transcription levels under dark condition.
在生物钟的生物振荡器中,CmFRQ为负调控蛋白,CmWC-1和CmWC-2为正调控蛋白。在对照组中,Cmwc-1和Cmwc-2前期的转录水平升高(培养的第6天)促进了Cmfrq转录水平的振荡上行(图3A,3B,3C)。
MG132为蛋白酶体抑制剂,能够使蛋白质泛素化降解受抑制。在MG132处理组中,第8-10天的Cmfrq转录水平急剧升高使细胞中聚集了一定量的未降解的CmFRQ,由此也使Cmwc-1和Cmwc-2在随后的一段时间内转录水平都不高(图3A,3B,3C)。
2.4 蓝光照射条件下10 μmol/L MG132对Cmfrq、Cmwc-1和Cmwc-2转录水平的影响
2.4.1 黑暗培养过程中2 h蓝光间断条件下,10 μmol/L MG132对Cmfrq、Cmwc-1和Cmwc-2转录水平的影响
蛹虫草接种在含有10 μmol/L MG132的PDA培养基上,黑暗条件下培养第5天的菌丝体进行蓝光照射2 h后第一次取样,剩余菌丝体样品重新置于黑暗中培养,每天同一时间取样,连续取样10 d,进行实时荧光PCR分析。其中Cmfrq转录水平受MG132的影响,在培养的前12 d大部分时间均低于对照组的转录水平,第12天以后MG132处理组的转录水平开始快速升高,对照组的转录水平急剧下降(图4A);Cmwc-1基因的转录水平在前12 d受MG132的影响不大,对照组和MG132处理组转录水平非常接近,但在第12天以后,对照组的转录水平急剧升高,而MG132处理组的转录水平变化不大(图4B);Cmwc-2在不加MG132的对照组中转录量起伏不大,但在加MG132的处理组中前10 d有一个转录量降低的过程,但在第10天以后转录量出现一个振荡上行的过程(图4C)。
图4
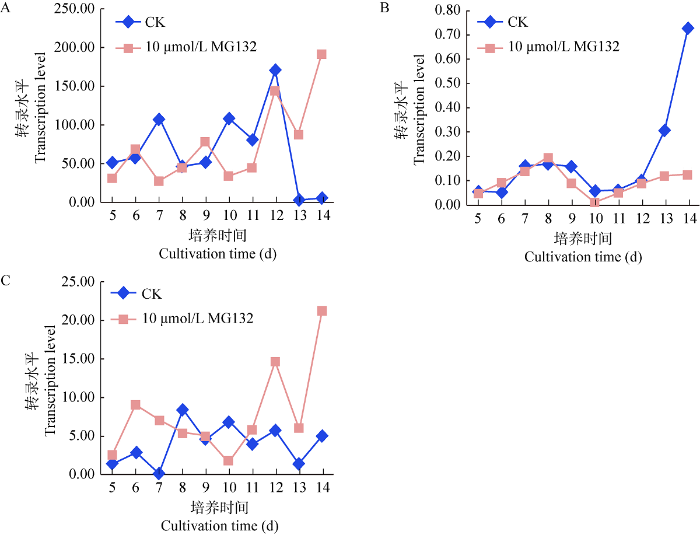
图4
蛹虫草菌丝体黑暗培养过程中2 h蓝光间断条件下,10 μmol/L MG132处理对Cmfrq (A)、Cmwc-1 (B)和Cmwc-2 (C)转录水平的影响
Fig. 4
Effects of 10 μmol/L MG132 treatment on Cmfrq (A), Cmwc-1 (B) and Cmwc-2 (C) transcription levels under dark culture condition of Cordyceps militaris mycelium with 2 h blue light interruption.
总体来看,不加MG132的对照组中,在培养的第12天开始Cmfrq转录水平急剧下降,Cmwc-1转录水平却急剧上升,出现了明显的生物振荡器中转录联动现象,但对Cmwc-2转录水平影响不大(图4A,4B,4C)。
在MG132处理组中,Cmfrq转录水平在后期出现一个上行趋势,Cmwc-2转录水平也是如此,但Cmwc-1转录水平却起伏不大(图4A,4B,4C)。由此推测,MG132处理组中Cmfrq、Cmwc-1和Cmwc-2基因转录失去了正调控和负调控的联动效应。
2.4.2 持续蓝光照射条件下10 μmol/L MG132对Cmfrq、Cmwc-1和Cmwc-2转录的影响
图5

图5
持续蓝光照射条件下10 μmol/L MG132对Cmfrq (A)、Cmwc-1 (B)和Cmwc-2 (C)转录水平的影响
Fig. 5
Under continuous blue light irradiation, effects of 10 μmol/L MG132 treatment on Cmfrq (A), Cmwc-1 (B) and Cmwc-2 (C) transcription levels.
在未加MG132处理的菌丝体中,Cmfrq的转录水平出现振荡走高,但在后期(蓝光连续培养的第13天)出现急剧下降,这与黑暗培养过程中2 h蓝光间断条件的情形类似。
2.5 不同蓝光光照条件下Cmfrq、Cmwc-1和Cmwc-2转录水平的比较
将结果2.3和2.4中黑暗培养、黑暗培养过程中蓝光间断2 h培养和持续蓝光照射培养的蛹虫草菌丝体中Cmfrq、Cmwc-1和Cmwc-2 3个基因,按照不加MG132的对照与加MG132的处理重新进行转录水平的分析比较。结果表明在不加MG132的对照中:(1)对3个基因转录水平进行比较,黑暗培养过程中蓝光间断照射2 h 和持续的蓝光照射情况下Cmfrq的转录水平基本都高于黑暗时的转录水平,但分别在12 d和13 d后均有快速下降(图6A),说明短期的蓝光照射和持续的蓝光照射在培养的前期会导致Cmfrq转录水平的提高,其中短期的蓝光照射更有利于Cmfrq转录水平的提高,在后期营养条件下降的情况下转录水平急剧下降,但对黑暗情况下Cmfrq转录水平的影响不大且转录水平低(图6A);(2) Cmwc-1的转录水平在这3种光照处理情况下的差异主要出现在前期和后期,其中持续蓝光照射(图6B)在前期(培养的第6天)转录水平突然升高,蓝光照射的前期对Cmwc-1的转录水平有较大影响;黑暗培养过程中蓝光间断照射2 h (图6B)在后期转录水平快速升高,表明蓝光照射后间隔较长时间才对Cmwc-1的转录水平有影响,且这种影响伴随着Cmfrq转录水平的急剧下降(图6A),同时也表明Cmfrq和Cmwc-1转录水平有一定的联动效果。黑暗条件下对Cmwc-1转录水平影响不大(图6B);(3) Cmwc-2的转录水平在这3种情况下的差异主要出现在黑暗处理的前期(图6C),即在第6天有一个大量的转录,其他两种情况无明显波动,且转录量很低;此后3种情况都维持低水平的转录,黑暗培养过程中蓝光中断照射2 h和持续蓝光照射情况下Cmwc-2平均转录水平均高于持续黑暗的转录水平(图6C)。
图6
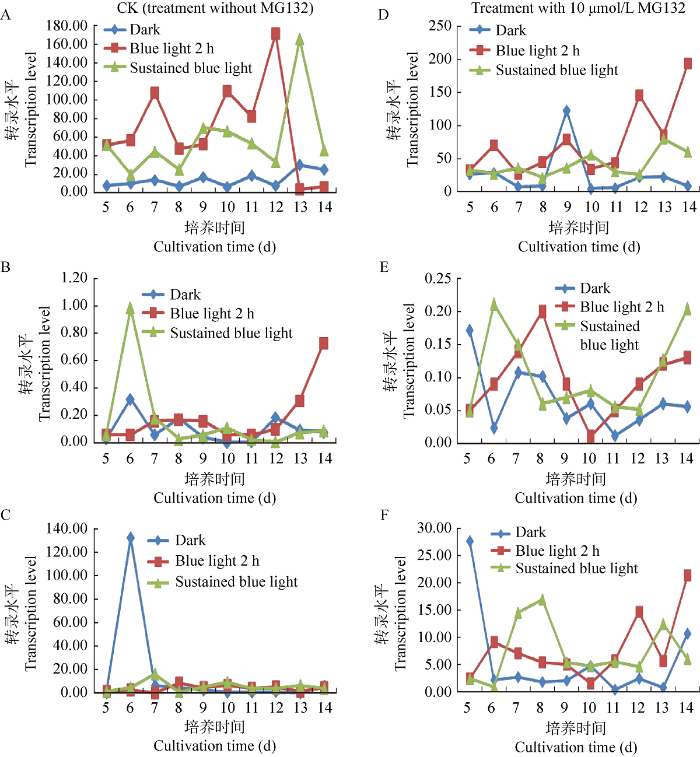
图6
有、无MG132处理条件下,不同蓝光光照时间对Cmfrq、Cmwc-1和Cmwc-2转录水平影响的分析
A:无MG132处理条件下,不同蓝光光照对Cmfrq转录水平影响的影响;B:无MG132处理条件下,不同蓝光光照对Cmwc-1转录水平影响的分析;C:无MG132处理条件下,不同蓝光光照对Cmwc-2转录水平影响的分析;D:有MG132处理条件下,不同蓝光光照对Cmfrq转录水平影响的影响;E:有MG132处理条件下,不同蓝光光照对Cmwc-1转录水平的影响;F:有MG132处理条件下,不同蓝光光照对Cmwc-2转录水平的影响
Fig. 6
Analyses of the influence of different duration of blue light irradiation on the transcription levels of Cmfrq, Cmwc-1, Cmwc-2 with or without MG132 treatment.
A: The effects of different duration of blue light irradiation on Cmfrq transcription level without MG132 treatment; B: The effects of different duration of blue light irradiation on Cmwc-1 transcription level without MG132 treatment; C: The effects of different duration of blue light irradiation on Cmwc-2 transcription level without MG132 treatment; D: The effects of different duration of blue light irradiation on Cmfrq transcription level with MG132 treatment; E: The effects of different duration of blue light irradiation on Cmwc-1 transcription level with MG132 treatment; F: The effects of different duration of blue light irradiation on Cmwc-2 transcription level with MG132 treatment.
在加10 μmol/L MG132的处理中:(1)总体来看,黑暗培养过程中蓝光间断照射2 h和持续的蓝光照射情况下Cmfrq的转录水平基本都高于黑暗时的转录水平,仅在第9天黑暗时的转录水平有个转录峰。此外,黑暗培养过程中蓝光间断照射2 h的处理,其Cmfrq的转录水平在后期振荡上行(图6D),这与无MG132处理的情况正好相反,表明短暂的蓝光照射和MG132处理可以导致后期Cmfrq转录水平的升高;(2) Cmwc-1的转录水平在这3种蓝光处理情况下,尽管有起伏变化,但总体转录水平均非常低,与对照比较并无急剧变化情况出现(图6E);(3) Cmwc-2的转录水平在黑暗培养的前期(第5天),或者更早时期可能有较高水平的转录,与不加MG132情形类似;此后3种情况都维持低水平转录,黑暗培养过程中蓝光间断照射2 h和持续蓝光照射情况下Cmwc-2平均转录水平要高于持续黑暗的转录水平(图6F),这与不加MG132的情形类似。
2.6 蓝光和黑暗交替条件下10 μmol/L MG132对蛹虫草Cmfrq、Cmwc-1和Cmwc-2基因在菌丝体中转录影响的分析
蛹虫草经过前5 d的黑暗培养后,在第6天早上6:00开始蓝光照射处理,之后每间隔2 h取一次样,在18:00关闭蓝光照射并进入黑暗培养条件下,每间隔2 h取一次样品,如此循环2 d进行蓝光和黑暗交替处理,一共24组样品,进行实时荧光PCR分析。结果表明,在第6天开始进入蓝光照射的12 h中,对照组中Cmfrq基因转录呈下降趋势,在进入黑暗以后的3-4 h达到转录量的低点,随后进行黑暗-蓝光-黑暗的交替中有一个起伏不大的波动;而MG132处理组,从第6天开始进入蓝光照射的12 h中,Cmfrq转录水平以4 h为周期,呈现一个较为剧烈的波动,在进入黑暗以后的6-8 h达到转录量的低点,随后进行黑暗-蓝光-黑暗的交替中有一个起伏不大的波动(图7A)。比较来看,对照和MG132处理Cmfrq转录水平的差异主要出现在前期12 h的蓝光照射,此后有、无MG132处理时Cmfrq转录水平差异不是很明显。
图7

图7
黑暗和蓝光照射交替条件下,10 μmol/L MG132对Cmfrq (A)、Cmwc-1 (B)和Cmwc-2 (C)转录水平的影响
Fig. 7
Effects of 10 μmol/L MG132 on the transcription levels of Cmfrq (A), Cmwc-1 (B) and Cmwc-2 (C) under alternating dark and blue light irradiation.
对3个基因的相关性进行分析,在不加MG132的情况下,Cmfrq基因在检测的2 d中均有转录,而Cmwc-1和Cmwc-2只在检测的第1个蓝光和黑暗周期中有转录,在第2个蓝光和黑暗周期中转录水平极低。这表明Cmfrq转录对Cmwc-1和Cmwc-2负调控作用主要体现在第2个蓝光和黑暗周期中。而在加MG132的情况下,Cmfrq、Cmwc-1和Cmwc-2这3个基因的相关性不明显。
3 讨论
真菌中研究生物钟生物振荡器主要成分之间的关系比较清楚的是在粗糙脉孢菌中(Heintzen & Liu 2007)。蛹虫草与粗糙脉孢菌同属子囊菌,彼此之间相关研究有一定的参照价值。但是,蛹虫草的3个基因Cmfrq、Cmwc-1和Cmwc-2与粗糙脉孢菌的3个基因frq (GenBank:U17073.1)、wc-1 (GenBank:X94300.2)和wc-2 (GenBank:Y09119.1)分别只有72.11%、71.23%和70.75%的同源性,势必导致蛹虫草中Cmfrq、Cmwc-1和Cmwc-2的转录与粗糙脉孢菌中frq、wc-1和wc-2的转录存在差异,这种差异必然体现在2种真菌的生物钟调控中。
粗糙脉孢菌作为模式真菌,其负反馈环中日夜调节机制中的正调控因子和负调控因子之间存在此消彼长的关联性。Cha et al. (2007)对粗糙脉孢菌中的相关研究进行了总结,在主观性早晨的持续黑暗中,WC-1和WC-2形成一种异二聚体复合物(D-WCC),该复合物与frq启动子中的时钟调控钟盒(Clock box或C box)结合,导致frq转录激活。frq mRNA在主观性白天达到峰值,FRQ蛋白量在4-6 h后达到峰值。合成FRQ蛋白后,它通过coiled-coil结构域与FRH形成复合物。在细胞核中,FFC抑制D-WCC的活性,导致frq mRNA水平降低;frq mRNA水平在主观性傍晚前后达到低谷。FRQ被磷酸化和去磷酸化后再经蛋白酶体系统泛素化和降解。当FRQ水平在主观性深夜下降到某个阈值以下时,D-WCC不再被FFC抑制,FRQ转录被重新激活以开始新的周期。显然,本文在蛹虫草的日夜交替的研究中,Cmfrq、Cmwc-1和Cmwc-2这3个基因的转录水平(图7)没有表现出与粗糙脉孢菌完全相同的变化。此外,在本文的研究中Cmwc-1转录水平一直比较低,成为了蛹虫草生物钟调节机制中的限定因素。由此推测,Cmwc-1转录水平低也导致与粗糙脉孢菌中负反馈环调控上的很大差异。
根据研究结果,在持续的黑暗培养过程中,加MG132和不加MG132对Cmfrq、Cmwc-1和Cmwc-2这3个基因的转录水平的影响有限,可以确定MG132作为蛋白酶体抑制剂在黑暗情况下处理蛹虫草菌丝体,在转录水平上不利于进行相关研究。在黑暗培养过程中2 h蓝光间断条件下,培养后期Cmfrq转录水平的急剧下降和Cmwc-1转录水平的急剧上升,反映出生物振荡器中CmFRQ和CmWC-1这2个成分分别为负调控和正调控成分;该条件下Cmfrq转录水平在后期的急剧下降与这个时候营养成分的下降有关,表明Cmfrq转录也受培养基中营养成分的调控,而MG132处理导致生物振荡器失效,使得Cmfrq转录对营养的改变不敏感。在持续的蓝光照射培养条件下,Cmfrq的转录水平在后期也出现急剧下降,表明也受后期营养成分下降影响,而MG132处理可以抑制Cmfrq转录;而有或没有MG132处理对Cmwc-1和Cmwc-2这2个基因的转录水平影响的差异有限,且这2种处理情况下这2个基因的转录趋势相近。持续的蓝光处理带来的结果无论是有或无MG132处理均没有反映出生物振荡器成分之间的联动关系,且Cmfrq转录对MG132处理更为敏感。在蓝光12 h和黑暗12 h的周期交替条件下,Cmfrq、Cmwc-1和Cmwc-2的转录水平并没有明显地反映出类似粗糙脉孢菌的生物节律性变化,推测蛹虫草的生物钟机制可能较为独特。
研究中发现无论是短暂的蓝光照射还是较长时间的蓝光照射均可导致Cmfrq转录水平大多数情况高于黑暗条件,这表明蓝光有助于Cmfrq转录水平的提高。正常情况下Cmfrq转录水平在蓝光条件下其后期的转录会受到营养条件下降的影响,而MG132可抑制这种对营养条件下降的敏感性。由此推测,蓝光有利于正调控因子的作用,进而促进Cmfrq转录;蛹虫草生物钟的正常运转也受到营养条件的影响。
上述结果仅是对Cmfrq、Cmwc-1和Cmwc-2转录水平的分析,考虑到生物振荡器的调控不仅在转录水平上,在翻译水平上、甚至在翻译后的修饰上也进行调控。因此相关研究还要进一步对Cmfrq、Cmwc-1和Cmwc-2这3个基因的转录和翻译产物进行细致的分析。
参考文献
Negative feedback defining a circadian clock: autoregulation of the clock gene frequency
The frequency (frq) locus of Neurospora crassa was originally identified in searches for loci encoding components of the circadian clock. The frq gene is now shown to encode a central component in a molecular feedback loop in which the product of frq negatively regulated its own transcript, which resulted in a daily oscillation in the amount of frq transcript. Rhythmic messenger RNA expression was essential for overt rhythmicity in the organism and no amount of constitutive expression rescued normal rhythmicity in frq loss-of-function mutants. Step reductions in the amount of FRQ-encoding transcript set the clock to a specific and predicted phase. These results establish frq as encoding a central component in a circadian oscillator.
Circadian rhythms from multiple oscillators: lessons from diverse organisms
The organization of biological activities into daily cycles is universal in organisms as diverse as cyanobacteria, fungi, algae, plants, flies, birds and man. Comparisons of circadian clocks in unicellular and multicellular organisms using molecular genetics and genomics have provided new insights into the mechanisms and complexity of clock systems. Whereas unicellular organisms require stand-alone clocks that can generate 24-hour rhythms for diverse processes, organisms with differentiated tissues can partition clock function to generate and coordinate different rhythms. In both cases, the temporal coordination of a multi-oscillator system is essential for producing robust circadian rhythms of gene expression and biological activity.
Posttranslational control of the Neurospora circadian clock
DOI:10.1101/sqb.2007.72.010
PMID:18419276
[本文引用: 1]

The eukaryotic circadian clocks are composed of autoregulatory circadian negative feedback loops that include both positive and negative elements. Investigations of the Neurospora circadian clock system have elucidated many of the basic mechanisms that underlie circadian rhythms, including negative feedback and light and temperature entrainment common to all eukaryotic clocks. The conservation of the posttranslational regulators in divergent circadian systems suggests that the processes mediating the modification and degradation of clock proteins may be the common foundation that allows the evolution of circadian clocks in eukaryotic systems. In this chapter, we summarize recent studies of the Neurospora circadian clock with emphasis on posttranslational regulation in the circadian negative feedback loop.
Functional conservation of light, oxygen, or voltage domains in light sensing
PAS domain-mediated WC-1/WC-2 interaction is essential for maintaining the steady-state level of WC-1 and the function of both proteins in circadian clock and light responses of Neurospora
DOI:10.1128/MCB.22.2.517-524.2002
PMID:11756547
[本文引用: 1]
In the frq-wc-based circadian feedback loops of Neurospora, two PAS domain-containing transcription factors, WHITE COLLAR-1 (WC-1) and WC-2, form heterodimeric complexes that activate the transcription of frequency (frq). FRQ serves two roles in these feedback loops: repressing its own transcription by interacting with the WC complex and positively upregulating the levels of WC-1 and WC-2 proteins. We report here that the steady-state level of WC-1 protein is independently regulated by both FRQ and WC-2 through different posttranscriptional mechanisms. The WC-1 level is extremely low in wc-2 knockout strains, and this low level of expression is independent of wc-1 transcription and FRQ protein expression. In addition, our data show that the PAS domain of WC-2 mediates the interactions of this protein with both WC-1 and FRQ in vivo. Such interactions are essential for maintaining the steady-state level of WC-1 and the proper function of WC-1 and WC-2 in circadian clock and light responses.
Coiled-coil domain-mediated FRQ-FRQ interaction is essential for its circadian clock function in Neurospora
DOI:10.1093/emboj/20.1.101 URL [本文引用: 1]
Interlocked feedback loops contribute to the robustness of the Neurospora circadian clock
WHITE COLLAR-1, a multifunctional Neurospora protein involved in the circadian feedback loops, light sensing, and transcription repression of wc-2
DOI:10.1074/jbc.M209592200 URL [本文引用: 1]
Neurospora wc-1 and wc-2: transcription, photoresponses, and the origins of circadian rhythmicity
Circadian rhythmicity is universally associated with the ability to perceive light, and the oscillators ("clocks") giving rise to these rhythms, which are feedback loops based on transcription and translation, are reset by light. Although such loops must contain elements of positive and negative regulation, the clock genes analyzed to date-frq in Neurospora and per and tim in Drosophila-are associated only with negative feedback and their biochemical functions are largely inferred. The white collar-1 and white collar-2 genes, both global regulators of photoresponses in Neurospora, encode DNA binding proteins that contain PAS domains and are believed to act as transcriptional activators. Data shown here suggest that wc-1 is a clock-associated gene and wc-2 is a clock component; both play essential roles in the assembly or operation of the Neurospora circadian oscillator. Thus DNA binding and transcriptional activation can now be associated with a clock gene that may provide a positive element in the feedback loop. In addition, similarities between the PAS-domain regions of molecules involved in light perception and circadian rhythmicity in several organisms suggest an evolutionary link between ancient photoreceptor proteins and more modern proteins required for circadian oscillation.
WC-2 mediates WC-1-FRQ interaction within the PAS protein-linked circadian feedback loop of Neurospora
DOI:10.1093/emboj/20.1.109 URL [本文引用: 1]
The Neurospora circadian system
The eukaryotic filamentous fungus Neurospora crassa has proven to be a durable and dependable model system for the analysis of the cellular and molecular bases of circadian rhythms. Pioneering genetic analyses identified clock genes, and beginning with the cloning of frequency (frq), work over the past 2 decades has revealed the molecular basis of a core circadian clock feedback loop that has illuminated our understanding of circadian oscillators in microbes, plants, and animals. In this transcription/translation-based feedback loop, a heterodimer of the White Collar-1 (WC-1) and WC-2 proteins acts both as the circadian photoreceptor and, in the dark, as a transcription factor that promotes the expression of the frq gene. FRQ dimerizes and feeds back to block the activity of its activators (making a negative feedback loop), as well as feeding forward to promote the synthesis of its activator, WC-1. Phosphorylation of FRQ by several kinases leads to its ubiquitination and turnover, releasing the WC-1/WC-2 dimer to reactivate frq expression and restart the circadian cycle. Light resetting of the clock can be understood through the rapid light induction of frq expression and temperature resetting through the influence of elevated temperatures in driving higher levels of FRQ. Several FRQ- and WC-independent, noncircadian FRQ-less oscillators (FLOs) have been described, each of which appears to regulate aspects of Neurospora growth or development. Overall, the FRQ/white collar complex feedback loop appears to coordinate the circadian system through its activity to regulate downstream-target clock-controlled genes, either directly or via regulation of driven FLOs.
Morphological observation of Cordyceps militaris treated with blue light and proteasome inhibitor MG132
White Collar-1, a circadian blue light photoreceptor, binding to the frequency promoter
In the fungus Neurospora crassa, the blue light photoreceptor(s) and signaling pathway(s) have not been identified. We examined light signaling by exploiting the light sensitivity of the Neurospora biological clock, specifically the rapid induction by light of the clock component frequency (frq). Light induction of frq is transcriptionally controlled and requires two cis-acting elements (LREs) in the frq promoter. Both LREs are bound by a White Collar-1 (WC-1)/White Collar-2 (WC-2)-containing complex (WCC), and light causes decreased mobility of the WCC bound to the LREs. The use of in vitro-translated WC-1 and WC-2 confirmed that WC-1, with flavin adenine dinucleotide as a cofactor, is the blue light photoreceptor that mediates light input to the circadian system through direct binding (with WC-2) to the frq promoter.
Rhythmic binding of a WHITE COLLAR-containing complex to the frequency promoter is inhibited by FREQUENCY
A PEST-like element in FREQUENCY determines the length of the ciradian period in Neurospora crassa
DOI:10.1093/emboj/20.24.7074 URL [本文引用: 2]
White collar-1, a DNA binding transcription factor and a light sensor
DOI:10.1126/science.1072795 URL [本文引用: 1]
Molecular mechanism of light responses in Neurospora: from light-induced transcription to photoadaptation
DOI:10.1101/gad.1369605 URL [本文引用: 1]
The Neurospora crassa circadian clock
Decoupling circadian clock protein turnover from circadian period determination
DOI:10.1126/science.1257277 URL [本文引用: 1]
The circadian pacemaker of mammals
DOI:10.1016/0968-0004(81)90006-2 URL [本文引用: 1]
Circadian rhythms in Neurospora crassa and other filamentous fungi
Circadian regulation of gene expression in animals
Circadian timekeepers reside in most body cells of Drosophila and mammals. The discovery of new clock genes suggests that circadian oscillations are generated by interconnected feedback loops employing transcriptional and post-translational mechanisms. In mammals, a master pacemaker localized in the suprachiasmatic nucleus synchronizes peripheral clocks via humoral cues. However, restricted feeding can uncouple peripheral oscillators from the suprachiasmatic pacemaker.
Role of a white collar-1-white collar-2 complex in blue-light signal transduction
DOI:10.1093/emboj/18.18.4961 URL [本文引用: 1]
Distribution of lysine pathways among fungi: evolutionary implications
DOI:10.1086/282338 URL [本文引用: 1]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}