


INTRODUCTION
Subtropical and tropical regions of China include the territory to the south of Qinling- Huaihe River Line. Wu (1979) classified this vast area into two floristic kingdoms: Holarctic and Paleotropic kingdoms. This area features a transition from temperate zone to the tropics with many endemic plant species. Representative ectomycorrhizal plants include Betulaceae, Fagaceae and Pinaceae. The high diversity of ectomycorrhizal trees suggests a rich ectomycorrhizal mycobiota that needs intensive investigation.
There have been many taxonomic studies of milk caps (Lactarius, Lactifluus and part of Multifurca), one of the most important ectomycorrhzial fungal groups, in subtropical and tropical China (Bi & Li 1987; Ying 1991; Wang et al. 2015a, 2018; Wang 2017, 2018). Among the 76 milk caps originally described from China, 56 are from the subtropical and tropical regions. These species include the most famous edible mushrooms such as L. vividus, Lf. tenuicystidiatus and Lf. tropicosinicus, endemic species specifically growing with vulnerable trees, L. guangdongensis, and species extending to countries of tropical Asia, e.g. L. brachycystidiatus, L. chichuensis and L. ambiguus (Wang et al. 2015a, 2015b; Wisitrassameewong et al. 2015; Wang 2017, 2018; Han et al. 2019).
From 2009 to 2021, the authors carried out a series of field investigations in subtropical and tropical China, focusing on the eastern (Anhui), central-southern (Hunan) and southwestern (Yunnan) provinces. In this paper, five new species and one species new to China are reported based on 48 specimens obtained from Anhui, Henan, Hunan, Jiangxi, Hainan and Yunnan. Morphological descriptions and DNA barcode ITS sequences for each species were provided.
1 MATERIALS AND METHODS
1.1 Morphological observation
For methods of morphological study, see Wang et al. (2021). Presence or absence of incrustations on hyphal surface was checked using distilled water. Voucher specimens were deposited in the Mycological Herbarium of Hunan Normal University, China (MHHNU), Cryptogamic section, Herbarium of Kunming Institute of Botany, Chinese Academy of Sciences (HKAS section, KUN) and Edible- medicinal Fungal Herbarium of Anhui Agricultural University, China (EFHAAU).
1.2 DNA extraction, PCR protocol, and sampling
DNA extractions and PCR protocols followed Wang et al. (2009) and Wang et al. (2012). Complete ITS regions (c. 700 bp) were amplified using primer pairs ITS1F+ITS4 or ITS5+ITS4 (White et al. 1990). For problematic samples whose PCR failed, primer pairs ITS1F+5.8S and ITS3+ITS4 were used to amplify the ITS-1 and ITS-2 regions separately. When sequences had heterozygous INDELs or ambiguous sites, samples were sequenced bidirectionally to make contigs of the amplified regions or verify the ambiguous sites. Flot et al.’s (2006) method was used to determine the INDEL heterozygosity and phase the sequences into two copies. Raw sequences were assembled with Sequencher v4.1.4 (Gene Codes Corporation, Ann Arbor), and consensus sequences were deposited in GenBank with accessions OL770135-OL770194 (Figs. 1 and 2). Both copies were submitted to GenBank for samples with two copies differing in INDELs, and the samples received two GenBank accessions.
Fig. 1
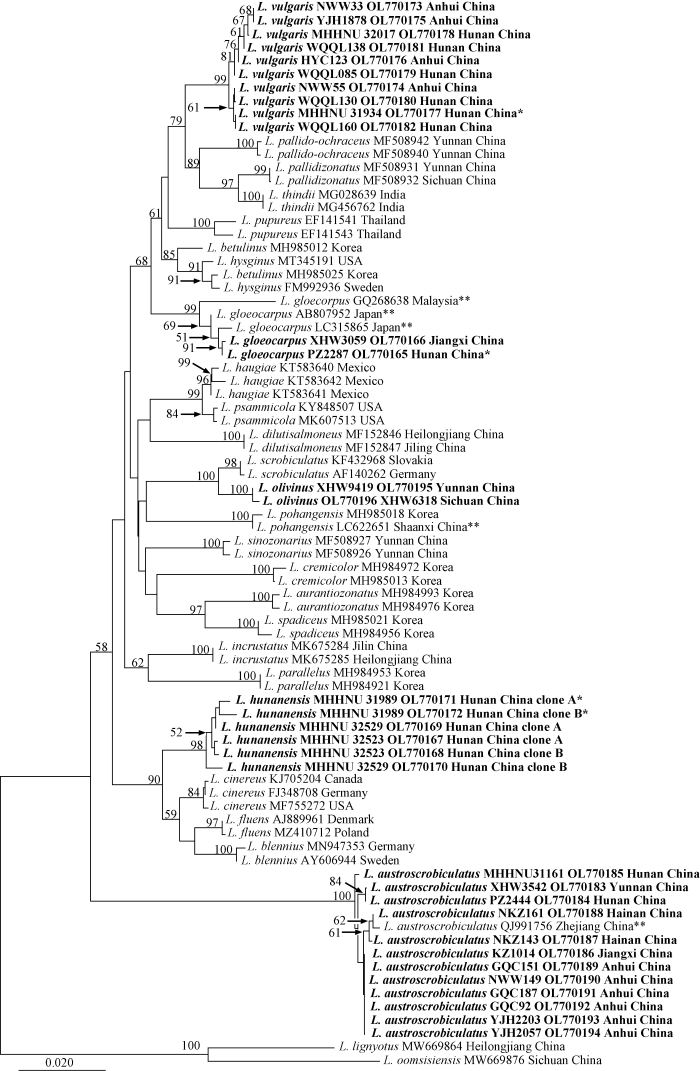
Fig. 1
Original tree generated by neighbor-joining (NJ) analysis of Lactarius subg.
Lactarius based on ITS sequences rooted with L. lignyotus and L. oomsisiensis. NJ bootstrap proportions higher than 50% are indicated above the branches. Sample names are presented in the order of species name, voucher specimen, GenBank accession, and geographical origin for the new species and new record. For the known species, voucher specimen information is left out. For Chinese samples, the province name is given. Type specimens are marked with an asterisk, and those from environmental samples are marked with two asterisks. Sequences generated in this study are in bold.
The generated ITS sequences (with the 18S and 28S ends trimmed) were submitted to BLASTn to find matches with high similarity (95%, a standard tolerant than OTUs) and query cover (>90%, covering the ITS-1, 5.8S, and ITS-2 regions). Sequences from both fruiting bodies and environmental samples were searched to determine the geographical distribution. For each species, matches with the same similarity and query cover were filtered, keeping up to three sequences representing different areas or/and showing clear genetic divergence. Using this strategy, 30 sequences of L. subg. Lactarius representing all 21 species retrieved were retained. To compare our new species with some Asian species with morphological resemblance, 47 sequences of L. aurantiozonatus, L. cremicolor, L. dilutisalmoneus, L. parallelus, L. pohangensis, L. sinozonarius, and L. thindii published by Wang (2017), Shi et al. (2018), Uniyal et al. (2018) and Lee et al. (2019) were added to the dataset. Four sequences of L. scrobiculatus and L. olivinus were included to test the relationship between these two species and L. austroscrobiculatus. To cover the known Asian species of subg. Plinthogalus, new sequences generated in this study were added to the ITS dataset of Wang et al. (2021), with some duplicate sequences of the same species removed. For the dataset of subg. Lactarius, L. lignyotus and L. oomsisiensis were selected as the outgroup taxa and for subg. Plinthogalus, L. torminosus and L. subdulcis were used to root the tree.
1.3 Phylogenetic analyses
Using the online version of the multiple sequence alignment program MAFFT v7 (Katoh & Toh 2008), alignments were made using the L-INS-I strategy. They were manually adjusted in BioEdit v.7.1.3.0 (Hall 1999). ITS datasets of subg. Lactarius and subg. Plinthogalus were made and analyzed separately. No partitioning was set for the ITS dataset, considering relatively low genetic divergence among the species. Due to the high similarity among sequences, we used the time-saving NJ method implemented in MEGA X (Kumar et al. 2018) to infer the phylogenetic relationships using the following settings: bootstrap method (with 1 000 replicates) as a test of phylogeny, p-distance as substitution model, transitions and transversions both included for substitutions, uniform rates among sites, the homogenous pattern among lineages and pairwise deletion for gaps treatment. An NJ bootstrap proportion (NJ-BP) ≥70% was considered significant support for a node to be monophyletic. Data matrices and trees are available at TreeBASE under S29118.
2 RESULTS
2.1 Phylogeny
The dataset of subg. Lactarius consists of 77 sequences of 25 species. Samples of each of the three new species were grouped into independent monophyletic clades with significant support (NJ-BP 91%-99%). The closest relatives of L. vulgaris are the Sino-Himalayan L. pallido-ochraceus, L. pallidizonatus and L. thindii (NJ-BP 79%). The two Chinese samples of L. gloeocarpus formed a clade with moderate support with two environmental samples from Japan (AB807952 and LC315865) (NJ-BP 69%) and this clade was sister to an environmental sequence from Malaysia (GQ268638) (NJ-BP 99%). Six sequences from three specimens of L. hunanensis formed a well-supported clade with temperate L. cinereus, L. fluens and L. blennius (NJ-BP 90%). Sequences of L. austroscrobiculatus formed a clade with the longest branch among all terminal clades. It is basal to all the other species, but this relationship lacked support (Fig. 2).
Fig. 2
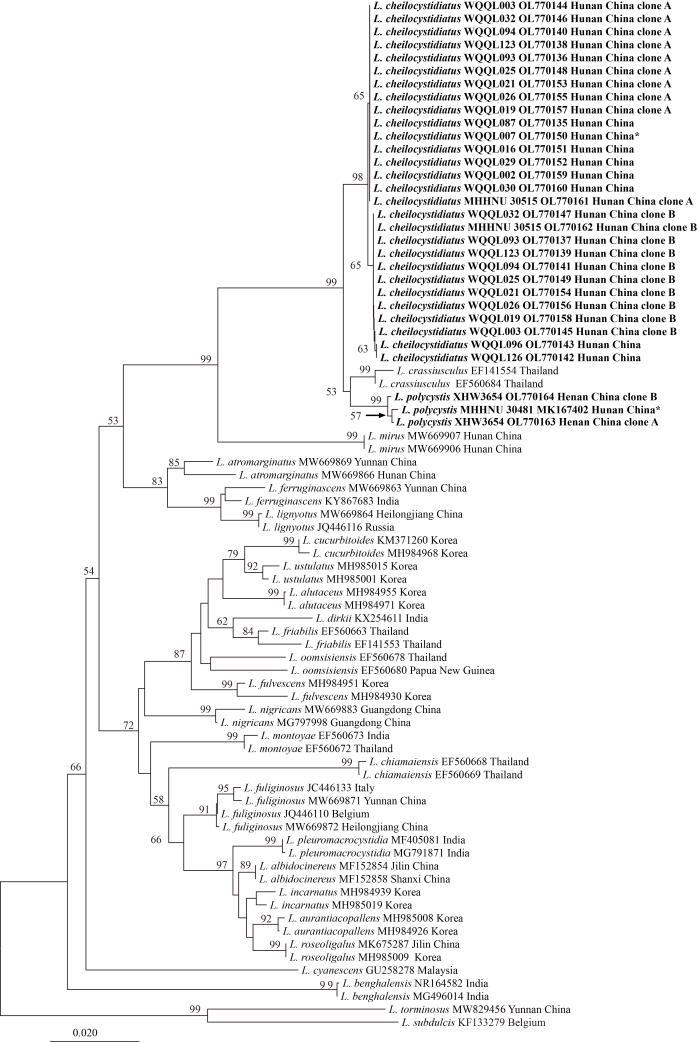
Fig. 2
Original tree generated by neighbor-joining (NJ) analysis of Lactarius subg.
Plinthogalus based on ITS sequences rooted with L. torminosus and L. subdulcis. NJ bootstrap proportions higher than 50% are indicated above the branches. Sample names are presented in the order of species name, voucher specimen, GenBank accession, and geographical origin for the new species and new record. For the known species, voucher specimen information is left out. For Chinese samples, the province name is given. Type specimens are marked with an asterisk. Sequences generated in this study are in bold.
The dataset of subg. Plinthogalus includes 78 nucleotide sequences. Samples of the two new species, L. cheilocystidiatus and L. polycystis, formed two well-supported clades (NJ-BP 97% and 98%), sister to Thai L. crassiusculus (NJ-BP 100%). The successive sister to the clade formed by the above three species is the newly described Chinese species L. mirus (NJ-BP 99%). Further relationships with other species were not resolved. All backbones lacked significant supports.
2.2 Taxonomy
Lactarius austroscrobiculatus Verbeken & E. Horak, Sydowia 53(2): 271. 2001. Fig. 3
Fig. 3
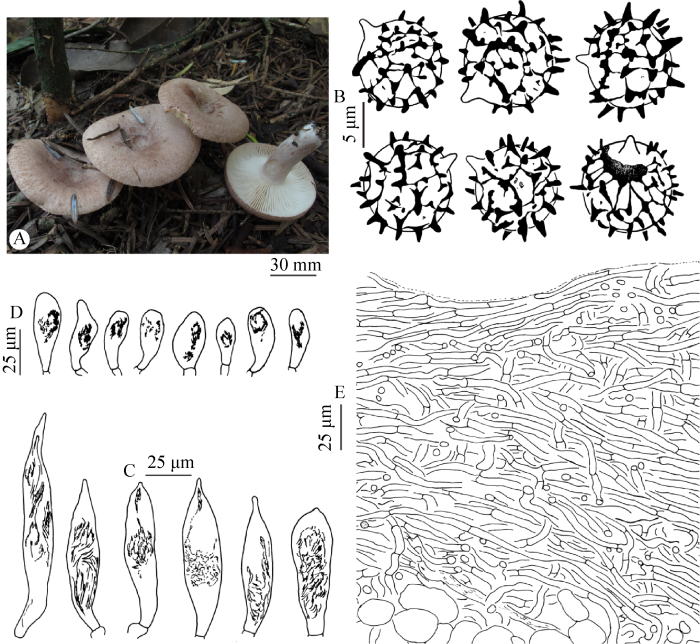
Fig. 3
Lactarius austroscrobiculatus (ZP-2444).
A: Basidiocarps; B: Basidiospores; C: Pleuromacrocystidia; D: Cheilomacrocystidia; E: Pileipellis.
DNA barcode ITS sequences: OL770183- OL770194.
Basidiocarps medium-sized, thick-fleshed. Pileus 60-90 mm diam., at first slightly depressed with a decurved margin, later becoming shallowly funnel-shaped with straight margin; surface dry or sticky, squamulose-felty to nearly glabrous, faintly to clearly zonate, hazel brown, often with purple, lilac or flesh-colored tinge. Lamellae straight to short decurrent, crowded, 5-6 mm broad, pale cream to pale buff. Stipe 30-90× 10-15 mm, cylindrical, equal or tapering downward; surface smooth, subviscid, scrobiculate or not, concolorous with or paler than the pileus surface. Context 3-4 mm thick, cream when young, later buff-yellow, yellowish-brown, or mahogany brown when bruised. Taste and smell unknown. Latex white, slowly changing to pinkish or brownish.
Basidiospores (7.5) 9.0-10.0-11.0 (12.0)× (7.0) 8.0-9.4-10.0 (11.0) µm [Q=(1.00) 1.02- 1.07-1.16 (1.28)] (240/12/11), subglobose, globose, rarely ellipsoid; ornamentation 1.0-2.5 (3.0) μm high, of acute warts and higher ridges connected by much lower and finer lines, forming a spinose-reticulate pattern; plage not or distally amyloid. Basidia 45-60× 12-17 µm, subclavate, 4-spored; sterigmata 8-15 µm long. Pleuromacrocystidia 54-128× 11-24 µm, scattered to common, emergent, fusiform, rarely clavate, often with a constricted or moniliform apex. Hymenophoral pseudocystidia rare, 5-6 µm broad, cylindrical, hyaline. Lamellar edge sterile; marginal cells 10-20×5-10 µm, cylindrical, clavate, hyaline; cheilomacrocystidia 25-43×9-17 µm, clavate, rare, with crystalline contents. Pileipellis an ixocutis, 150-250 μm thick, of loosely or compactly interwoven hyphae; hyphae 2-6 μm broad, slightly gelatinized, some shriveled, hyaline. Stipitipellis an ixocutis, 60-100 μm thick; hyphae 2-4 μm broad, hyaline, gelatinized. Hymenophoral trama with abundant golden brownish lactifers. Trama of pileus and stipe with abundant rosettes. Clamp connections absent.
Habitat and distribution: in subtropical and tropical broad-leaved forests or mixed forests with Abies trees, gregarious. July to September. Central and southern China (Anhui, Hainan, Hunan, Jiangxi, Yunnan and Zhejiang).
Specimens examined: CHINA. Anhui: Huangshan City, Huangshan District, Ciguangge, N30°10ʹ26.35ʺ E118°16ʹ65.97ʺ, alt. 727 m, 11 Sept. 2018, G.Q. Chu, CGQ92 (EFHAAU 1306); Huangshan City, Huangshan scenic area, road from Yongdingqiao to Yunmenfeng, N30°07ʹ55.39ʺ E118°07ʹ36.63ʺ, alt. 632 m, 13 Sept. 2018, Y.J. Hao, HYJ2057 (EFHAAU 655); Huangshan City, Huangshan District, Huangshan Reservoir, N30°11ʹ16.04ʺ E118°15ʹ78.28ʺ, alt. 861 m, 15 Sept. 2018, G.Q. Chu, CGQ151 (EFHAAU 1365); Huangshan City, Huangshan Biodiversity Monitoring Site, 14 Sept. 2018, N.W. Wang, WNW149 (EFHAAU 1718); Shitai County, Guniujiang Nature Reserve, N30°06ʹ38.06ʺ E117°48ʹ03.31ʺ, alt. 421 m, 19 Sept. 2018, G.Q. Chu, CGQ187 (EFHAAU 1401); Shitai County, Guniujiang Nature Reserve, Shuanglonggou, Huochawu, N30°03ʹ49.83ʺ E117°28ʹ49.28ʺ, alt. 432 m, 19 Sept. 2018, Y.J. Hao, HYJ2203 (EFHAAU 801). Hainan: Qiongzhong County, Limushan Nature Reserve, 13 May 2009, N.K. Zeng, 147 (KUN-HKAS 86963), 161 (KUN-HKAS 60419). Hunan: Yizhang County, Mangshan Forest Park, Zeziping, 14 Sept. 2016, P. Zhang, ZP-2444 (MHHNU); Guidong County, Bamianshan, 5 Jul. 2018, Z.H. Chen, MHHNU 31161 (MHHNU). Jiangxi: Jingan County, Sanzhualun, 3 Sept. 2017, K. Zhao, ZK1014 (KUN-HKAS 122609). Yunnan: Pingbian County, Daweishan Nature Reserve, Shuiweicheng, alt. 2 000 m, 9 Jul. 2012, X.H. Wang, 3542 (KUN-HKAS 76076).
Note: Lactarius austroscrobiculatus was originally described from Java, Indonesia, growing in montane rainforest dominated by Castanopsis and Quercus trees (Verbeken et al. 2001). This is the first record of the species in China. The diagnostic characteristics of this species are the typical hazel brown pileus with lilac to fleshed-colored tinge, big globose spores with dense spinose-reticulate ornamentation and large pleuromacrocystidia. The general feature of the Chinese specimens examined, especially that of ZP2444, matches up with these core features. Latex of the Chinese specimens does not change or slightly discolors pinkish (in KUN-HKAS 76076). This does not seem to deviate from the yellowing latex of the holotype because the “yellowing” reaction was observed when mounted with KOH (Verbeken et al. 2001). The pileus can be faintly zonate (in KUN- HKAS 60419, ZP2444, and KUN-HKAS 76076) to clearly zonate (in KUN-HKAS 122609). Scrobicules were absent in some specimens (KUN-HKAS 76076 and 60419).
This species has the furthest genetic distance from other species of subg. Lactarius (as a long branch in Fig. 1). Its phylogenetic position needs further study. In GenBank, the closest hit is an environmental sequence (QJ991756) from Gutianshan, Zhejiang, China, with a similarity of 98.95%. The finding of this species in southern China will help understand the mycobiotic connection between China and tropical Asia. There is no report on the edibility of this species.
Lactarius cheilocystidiatus X.H. Wang, W.Q. Qin & Fang Wu sp. nov.Fig. 4
Fig. 4
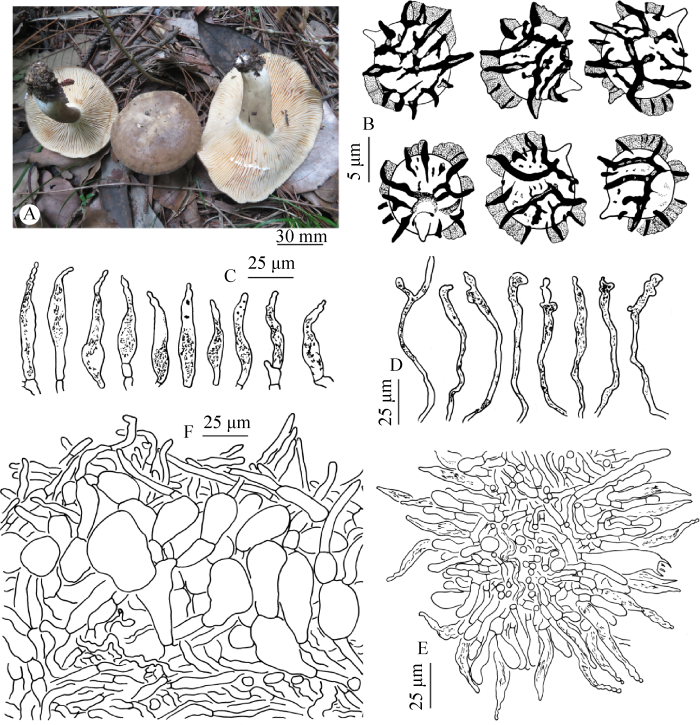
Fig. 4
Lactarius cheilocystidiatus (KUN-HKAS 110925, holotype).
A: Basidiocarps; B: Basidiospores; C: Pleuromacrocystidia; D: Hymenophoral pseudocystidia; E: Lamellar edge; F: Pileipellis.
MycoBank MB 842230
DNA Barcode ITS sequences: OL770150 (holotype), OL770135-OL770162
Holotype: CHINA. Hunan: Zhangjiajie City, Yongding District, Zhangjiajie Campus, Jishou University, N29°8ʹ32.62ʹʹ E110°27ʹ38.04ʹʹ, alt. 230 m, in forests dominated by Pinus massoniana, Quercus aliena var. acutiserrata and Q. serrata, 17 Jun. 2020, W.Q. Qin, L007 (KUN-HKAS 110925)
Etymology: refers to having abundant cheilomacrocystidia.
Basidiocarps medium-sized, thick-fleshed. Pileus 30-100 mm diam., hemispherical when young, convex with depressed center or shallowly funnel-shaped when mature, margin even or crenate, sometimes rugulose to finely radially folded; surface dry, shiny when wet, pruinose-subvelvety, grayish-brown to pale ochraceous brown, often with whitish spots. Lamellae adnate to short decurrent, crowded when young, subcrowded when fully mature, 2-6 mm broad, white when young, later brownish, warm buff when fully mature, stained flesh pink and finally dark brown where bruised. Stipe 40-75×6-12 mm, solid, subcylindrical or slightly enlarged downward, sometimes compressed; surface dry, smooth, grayish white to grayish brown, often paler than the pileus, apex white, base with white tomentum. Context 2-5 mm thick, white, turning flesh pink when cut, finally changing to brownish. Taste and smell unknown. Latex white, turning flesh pinkish slowly, becoming dark brown on lamellae. Spore print ochraceous.
Basidiospores (7.0) 8.0-8.7-9.5 (10.0)× (6.0) 7.0-8.1-9.0 µm [Q=(1.00) 1.02-1.08- 1.13 (1.29)] (140/7/6), globose, subglobose, very rarely ellipsoid; ornamentation up to 3.0 μm high, mostly 1.5-2.5 μm high, of acute heavy ridges, wings and small isolated warts, ridges branched, often less amyloid at the top rendering a split appearance; plage distally amyloid, sometimes not amyloid. Basidia 42-64×11-16 µm, subclavate, 4-spored; sterigmata 4-7 µm long. Pleuromacrocystidia 42-65×7-12 µm, only present near the lamellar edge, fusiform, apex blunt or moniliform, with granular contents. Lamellar edge fertile; cheilomacrocystidia 47-90× 6-13 µm, abundant, fusiform to sunfusiform, with granular or crystalline contents, apex often pointed and moniliform. Hymenophoral pseudocystidia common to moderately abundant, 2-6 μm broad at the base, with apex enlarged up to 6-12 μm broad, tortuous, apex often forking or branching, thin-walled, with strongly refractive yellowish contents. Pileipellis a trichoepithelium, 80-150 μm thick, with yellowish brown pigmentation; suprapellis 50-70 μm thick, terminal elements 15-60×3-7 μm, equal or slightly tapering, rarely narrowly clavate; subpellis 30-50 μm thick, pseudoparenchymatous, cells subglobose, globose, some hyphoid, 12-30 μm diam., slightly gelatinized in some individuals. Stipitipellis a trichoderm, 60-100 μm thick; terminal hyphae hyaline to pale yellowish brown, slightly gelatinized, 24-57×3-5 μm. Hymenophoral trama with abundant lactifers and hyphoid elements. Trama of pileus and stipe with abundant rosettes. Clamp connections absent.
Habitat and distribution: in subtropical forests dominated by Pinus massoniana, Quercus serrata, Q. fabri and Q. aliena var. acutiserrata, sometimes as well as Platycarya strobilacea, solitary or gregarious. June to September. Only known from Hunan.
Additional specimens examined: CHINA. Hunan: Shimen County, Huping Mountain Nature Reserve, Changlin, 18 Sept. 2009, Z.H. Chen, MHHNU 30515 (MHHNU); Zhangjiajie City, Yongding District, Zhangjiajie Campus, Jishou University, N29°8ʹ32.62ʹʹ E110°27ʹ38.04ʹʹ, alt. 230 m, in forests dominated by P. massoniana, Q. aliena var. acutiserrata and Q. serrata, alt. 230 m, 14 Jun. 2020, W.Q. Qin, L001 (KUN-HKAS 110919); ibid., 3 Jul. 2020, W.Q. Qin, L012 (KUN-HKAS 110930); ibid., 10 Jul. 2020, W.Q. Qin, L002 (KUN- HKAS 121207); ibid., 9 Jul. 2020, W.Q. Qin, L003 (KUN-HKAS 121210); ibid., 16 Sept. 2020, W.Q. Qin, L019 (KUN-HKAS 110942); ibid., 17 Sept. 2020, W.Q. Qin, L021 (KUN-HKAS 110944); ibid., 20 Sept. 2020, W.Q. Qin, L025 (KUN-HKAS 110949); ibid., 21 Sept. 2020, W.Q. Qin, L026 (KUN-HKAS 110950); ibid., 26 Sept. 2020, W.Q. Qin, L030 (KUN-HKAS 110954), L032 (KUN-HKAS 110956); ibid., 25 Aug. 2021, W.Q. Qin, L087 (KUN-HKAS 121338); ibid., 2 Sept. 2021, W.Q. Qin, L092 (KUN-HKAS 121343); ibid., 3 Sept. 2021, W.Q. Qin, L093 (KUN-HKAS 121344), L094 (KUN-HKAS 121345); ibid., 6 Sept. 2021, W.Q. Qin, L123 (KUN-HKAS 121374); ibid., N29°8ʹ33.25ʹʹ E110°27ʹ37.65ʹʹ, alt. 257 m, in forests dominated by Q. serrata, Q. aliena var. acutiserrata, and P. strobilacea, 3 Sept. 2021, W.Q. Qin, L096 (KUN-HKAS 121347); ibid., 6 Sept. 2021, W.Q. Qin, L126 (KUN-HKAS 121377).
Note: the most distinctive features of L. cheilocystidiatus are the grayish brown pileus, spores with prominent ornamentation and numerous macrocystidia that are mostly at the lamellar edge. This new species is close to L. crassiusculus and shares grayish brown pileus, crowded lamellae, white latex and presence of macrocystidia with the latter (Le et al. 2007). Unlike this new species, L. crassiusculus has numerous pleuromacrocystidia.
Species with macrocystidia are quite rare in subg. Plinthogalus (Stubbe et al. 2008). Besides L. crassiusculus (Le et al. 2007), three other species, L. papillatus (from Malaysia, Stubbe et al. 2008), L. lignyotus (from Europe, Heilmann-Clausen et al. 1998) and L. pleuromacrocystidia (from India, Uniyal et al. 2019) have macrocystidia. Differing from the new taxon, L. papillatus has a more yellowish pileus and distant lamellae. The cystidia of the latter two species do not have any contents, therefore, whether they represent real macrocystidia needs further consideration. In subg. Plinthogalus, L. atromarginus has been reported to be poisonous, causing gastroenteritis in China (Li et al. 2021). Considering the close relationships among species within subg. Plinthogatlus, we do not recommend L. cheilocystidiatus as edible mushroom.
Lactarius vulgaris Fang Wu, X.H. Wang, W.Q. Qin, Y.J. Hao & Z.H. Chen sp. nov.Fig. 5
Fig. 5
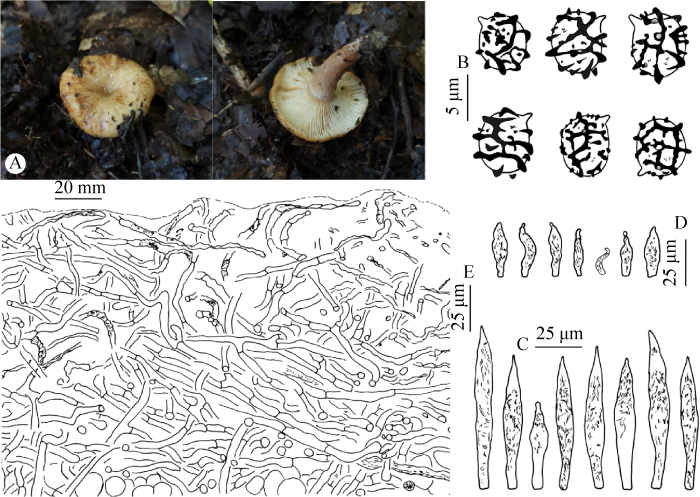
Fig. 5
Lactarius vulgaris (MHHNU 31934, holotype).
A: Basidiocarps; B: Basidiospores; C: Pleuromacrocystidia; D: Cheilomacrocystidia; E: Pileipellis.
MycoBank MB 842233
DNA barcode ITS sequences: OL770177 (holotype), OL770173-OL770182.
Holotype: CHINA. Hunan: Sangzhi County, Badagong Mountain Nature Reserve, in forest dominated with Fagus lucida and Betula insignis, 27 Jul. 2020, Z.H. Chen, MHHNU 31934 (MHHNU!)
Etymology: means “common”, referring to the frequent occurrence of the species in central-eastern China.
Basidiocarps medium-sized to large. Pileus 30-90 mm diam., convex when young, becoming broadly convex to plane with a depressed center, or shallowly funnel-shaped, margin decurved when young, becoming expanded and wavy when mature; surface glabrous, viscid, shiny, hygrophanous, faintly zonate, cream yellow, clay-buff to cinnamon, center brown, rarely with scrobicules along the margin. Lamellae adnate to decurrent, crowded, 1-2 mm wide, whitish to pale cream buff, turning pale yellowish brown when bruised. Stipe 30-80×10-20 mm, often eccentrical, cylindrical, spindle-shaped, or tapering downward; surface smooth, dry to slightly greasy, pale cream to pale pinkish buff, sometimes with big scrobicules. Context 2-3 mm thick, whitish to pale cream. Taste and smell unknown. Latex copious, white, unchanging.
Basidiospores (4.5) 5.5-6.3-7.0 (8.0)× 4.5-5.4-6.0 (7.0) µm [Q=(1.03) 1.07-1.16- 1.25 (1.36)] (160/8/7), subglobose to ellipsoid; ornamentation up to 1.0 µm high, mostly 0.3-0.7 µm high, of blunt warts and ridges forming a partial to nearly complete reticulum, isolated warts and irregular short ridges common; plage not amyloid. Basidia 25-48× 7-12 µm, subclavate, cylindrical, 4-spored, sterigmata 5-8 µm long. Pleuromacrocystidia (43) 55-80×(7) 9-15 µm, scattered, emergent, fusiform with an acute apex, upper part with dense granular or needle-like contents, contents colorless or pale yellowish brown, strongly refractive, apex often constricted to moniliform. Lamellar edge sterile; marginal cells 10-20×5-8 µm, clavate, capitate, hyaline; cheilomacrocystidia 30-45×6-10 µm, scattered, subfusiform to fusiform, apex blunt or pointed. Hymenophoral pseudocystidia rare, 3-4 µm broad, hyaline. Pileipellis an ixocutis to ixolattice, 120-230 µm thick, of loosely interwoven hyphae; hyphae 2-5 µm broad, hyaline to pale yellowish brown, mostly strongly shriveled, slightly gelatinized. Stipitipellis an ixocutis or ixotrichoderm, 25-100 µm thick; hyphae 2-5 µm broad, those in subpellis slightly gelatinized, loosely or densely interwoven, hyaline, locally with yellowish-brown lactifers. Hymenophoral hyphae 3-10 µm broad and abundant hyaline or light yellowish brown lactifers. Trama of pileus and stipe with abundant rosettes. Clamp connections absent.
Habitat, habit, and distribution: in forests dominated by fagaceous trees and sometimes birches and hornbeams, solitary or 2-3 individuals growing together. July to September. Central-southern China (Anhui and Hunan).
Additional specimens examined: CHINA. Anhui: Huangshan City, Huangshan, road from Ciguangge to Tianhai, N30°07′21.08″ E118°10′12.84″, alt. 1 516 m, 17 Jul. 2018,H.Y. Chen, CHY123 (KUN-HKAS 122608, KUN); Yuexi County, Yaoluoping Nature Reserve, Qilingou, 21 Jul. 2018, N.W. Wang, WNW33 (EFHAAU 1602); ibid., 23 Jul. 2018, N.W. Wang, WNW55 (EFHAAU 1624); Yuexi County, Yaoluoping Nature Reserve, Yuexingwan, N31°04ʹ34.94ʺ E116°10ʹ02.55ʺ, alt. 1 077 m, 27 Jul. 2018, Y.J. Hao, HYJ1878 (EFHAAU 476). Hunan: Sangzhi County, Badagong Mountain Nature Reserve, in forest dominated with Fagus lucida and Betula insignis, 29 Jul. 2020, Z.H. Chen, MHHNU 32017 (MHHNU); Zhangjiajie City, Yongding District, Tianmen Mountain, in forest dominated with Cyclobalanopsis multinervis, Quercus engleriana and F. lucida, N29°03′1″ E110°28′44″, alt. 1 470 m, 18 Jul. 2021, W.Q. Qin, L085 (KUN-HKAS 121336); ibid., in forest dominated with Q. engleriana, C. multinervis, F. lucida, and Carpinus fargesiana, N29°3ʹ6.44ʺ E110°29ʹ1.34ʺ, alt. 1 380 m, 9 Sept. 2021, W.Q. Qin, L138 (KUN-HKAS 121427); ibid. in forests dominated by C. multinervis and Lithocarpus cleistocarpus, N29°03′4.60″ E110°28′57.46″, alt. 1 381 m, 9 Sept. 2021, W.Q. Qin, L130 (KUN-HKAS 121419); in forest dominated with F. lucida and Cyclobalanopsis multinervis, N29°3ʹ6.01ʺ E 110°29ʹ2.54ʺ, alt. 1 357 m, 9 Sept. 2021, W.Q. Qin, L141 (KUN-HKAS 121430); ibid., in forest dominated with Cyclobalanopsis multinervis and Carpinus fargesiana, N 29°2ʹ52.61ʺ E110°28ʹ31.40ʺ, alt. 1 454 m, 12 Sept. 2021, L160 (KUN-HKAS 121449).
Note: The key characters of this new species are the faintly zonate viscid pileus with ochraceus tinge, small reticulate spores and large emergent macrocystidia. It is closely related to L. pallido-ochraceus, L. pallidizonatus and L. thindii. Lactarius vulgaris has the smallest and the least reticulate spores among the four species (L. pallido-ochraceous: 6.0-7.5×5.0-6.5 µm; L. pallidizonatus: 7.0-9.5×6.0-7.5 µm (Wang 2017); L. thindii: 6.0-7.0×5.0-6.0 µm (Uniyal et al. 2018). The pileus of this species varies from pale cream yellow, pale ochraceous (sometimes with olivaceous tinge) to pale flesh brown. The center is invariably darker than the margin, which resembles L. pohangensis. Lactarius pohangensis, however, has much smaller pleurocystidia (35.5-59×7.5-10.5 µm) and less reticulate spores (Lee et al. 2019). There is no report on the edibility of this new species, but Li et al. (2022) reported that its close relative L. purpureus stimulates the parasympathetic nervous system. Therefore we do not recommend this species as edible mushroom.
Lactarius gloeocarpus Fang Wu, X.H. Wang & Z.H. Chen sp. nov.Fig. 6
Fig. 6

Fig. 6
Lactarius gloeocarpus (ZP-2287, holotype).
A: Basidiocarps; B: Basidiospores; C: Pleuromacrocystidia; D: Cheilomacrocystidia; E: Pileipellis.
MycoBank MB 842236
DNA Barcode ITS sequences: OL770165 (holotype), OL770166
Holotype: CHINA. Hunan: Yizhang County, Mangshan Forest Park, Jiangjunzhai, alt. 1 400 m, 27 Jul. 2016, P. Zhang, ZP-2287 (MHHNU)
Etymology: the viscid pileus and stipe.
Basidiocarps medium-sized to large. Pileus 50-80 mm diam., convex with a decurved margin when young, later plano-convex with a depressed center, becoming shallowly funnel- shaped with a slightly wavy margin when mature; surface strongly viscid, not to faintly zonate, glabrous, pale yellow to light yellow, sometimes with greenish tinge, pale yellowish-brown at the center. Lamellae adnate to short decurrent, 4 mm broad, crowded, pale yellow when young, with more ochraceous tinge when mature, discoloring brownish when bruised. Stipe 30-60×12- 15 mm, equal or slightly spindle-shaped, yellowish white to pale yellow, very sticky, with conspicuous ochraceous scrobicules mostly at the lower part of the stipe. Context 3 mm thick, yellowish-white. Taste and smell unknown. Latex white, unchanging, taste immediately very bitter and sharp.
Basidiospores 6.0-7.2-8.0 (8.5)×5.0- 6.3-7.0 µm [Q=(1.03) 1.04-1.14-1.25 (1.37)] (60/3/2), subglobose to ellipsoid; ornamentation up to 1.3 μm high, mostly 0.3-1.0 μm high, of clean, heavy and blunt ridges forming an almost complete to complete reticulum, isolated warts present; plage not amyloid. Basidia 30-40×7-10 µm, subclavate, 4-spored; sterigmata 4-5 µm long. Pleuromacrocystidia 40-75×8-18 µm, common, emergent, projecting 20-40 µm beyond the basidia layer, subfusiform or ventricose with a pointing and often moniliform apex, with yellowish granular contents. Lamellar edge fertile; marginal cells 12-16×6-9 µm, clavate, colorless; cheilomacrocystidia 25-37×7-9 µm, scattered, subfusiform, fusiform, apex pointed, with yellowish needle-like contents. Hymenophoral pseudocystidia rare, sometimes slightly emergent, 3-4 µm broad, subcylindrical, hyaline, with strongly refractive contents. Pileipellis a typical ixotrichoderm, 150-300 μm thick (including the slime layer 40-150 μm thick), of loosely interwoven hyphae; hyphae ascending, 2-5 µm broad, terminal elements and several sections below often broader and shorter, forming a chain of short cells, apex blunt, some shriveled, not gelatinized. Stipitipellis an ixocutis, 40-70 µm thick, of loosely interwoven hyphae; hyphae 2-3 µm broad, hyaline. Hymenophoral trama with abundant pale yellowish brown lactifers. Trama of pileus and stipe with abundant rosettes. Clamp connections absent.
Habitat and distribution: in forests dominated by Pinus taiwanensis and fagaceous trees, 2-3 individuals together, July to September. Central-southern China (Hunan and Jiangxi) and Japan.
Additional specimens examined: CHINA. Jiangxi: Jiujiang City, Lushan, Lulin lakeside, N29°33ʹ13.32ʺ E115°58ʹ42.41ʺ, alt. 1 040 m, 1 Sept. 2011, X.H. Wang, 3059 (KUN-HKAS 73602).
Note: the pale yellow basidiocarps, the pileipellis a typical ixotrichoderm and the spores with a clean reticulum are diagnostic features of this new species. Such pileipellis is very often found in temperate and subalpine species, such as L. albocarneus (Heilmann- Clausen et al. 1998), L. caespitosus (Hesler & Smith 1979) and L. castaneus (Wang 2007), but not recorded in subtropical or tropical species. These temperate/subalpine species all have larger spores and different colors. Two environmental sequences from Japan, AB807952 (as “Lactarius sp. 6” in Murata et al. 2013) and LC315865 (as “Lactarius sp.4” in Murata et al. 2017) suggest the species also forms ectomycorrhizae with Abies firma and Tsuga sieboldii. The latex of this species is very bitter and sharp and we regard it as an inedible species.
Lactarius hunanensis Fang Wu, Z.H. Chen & X.H. Wang sp. nov.Fig. 7
Fig. 7

Fig. 7
Lactarius hunanensis (all from MHHNU 31989, holotype except B from MHHNU 32523).
A, B: Basidiocarps; C: Basidiospores; D: Pleuromacrocystidia; E: Cheilomacrocystidia; F: Pileipellis.
MycoBank MB 842237
DNA barcode ITS sequences: OL770171 and OL770172 (holotype), OL770167-OL770170.
Holotype: CHINA. Hunan: Sangzhi County, Badagong Mountain Nature Reserve, in forest dominated by Fagus lucida and Betula insignis, 27 Jul. 2020, Z.H. Chen, MHHNU 31989 (MHHNU!).
Etymology: the type locality, Hunan Province.
Basidiocarps small to medium-sized, slender. Pileus 20-60 mm diam., plano-convex with a decurved margin, margin±wavy; surface strongly viscid, pale grayish cream to grayish yellow or even olive brown, darker at the center, with a conspicuous green tinge when dry. Lamellae subdecurrent to decurrent, crowded, narrow, 1 mm broad, whitish, cream-white to yellowish cream, pale ochraceous when mature, faintly stained orange-brown. Stipe 36-80×7-10 mm, cylindrical or gradually enlarged toward the base, hollow; surface greasy, pale cream to pale grayish-brown, with greenish tinge when dry, sometimes with scrobicules. Context 1-2 mm thick, whitish cream. Taste and smell unknown. Latex whitish, unchanging.
Basidiospores (5.5) 6.0-7.0-8.0 (9.0)× 5.0-5.9-7.0 (8.0) µm [Q=(1.03) 1.13-1.19- 1.31 (1.40)] (80/4/3), ellipsoid; ornamentation 0.3-1.0 μm high, of ridges and isolated irregular warts aligned in a zebroid pattern, some ridges branched but never forming closed meshes; plage not amyloid. Basidia 4-spored, 25-35×7-11 µm, subclavate, sterigmata 4-6 µm long. Pleuromacrocystidia common, 38-75×5-11 µm, the base from the same depth as the basidia, narrowly fusiform, lanceolate, often with an acute and moniliform apex, with pale yellowish granular or crystalline contents. Hymenophoral pseudocystidia scattered, 2-4 µm broad, subcylindrical, sometimes bifurcate, hyaline. Lamellar edge fertile; marginal cells clavate, hyaline, 10-18 (25)×4-7 µm; cheilomacrocystidia scattered to common, 18-35 (47)×5-9 µm, subfusiform, with pale yellowish granular contents. Pileipellis an ixolattice, 70-200 μm thick in water, 150-280 μm thick in KOH, of loosely interwoven hyphae; hyphae 3-5 µm broad, occasionally inflated to 3-5 µm, not gelatinized, with numerous olivaceous incrustations (when observed in water), incrustations unevenly distributed, more common on hyphae beneath the subpellis, soon dissolved in KOH, terminal elements irregularly projecting outwards, apex blunt, 3-6 μm broad. Stipitipellis an ixocutis or ixolattice, 30-50 μm thick in water, 100-120 μm thick in KOH, of loosely interwoven hyphae; hyphae 2-5 μm broad, slightly gelatinized in subpellis, some shriveled, occasionally inflated to 10 μm, with olivaceous incrustations. Hymenophoral trama with abundant yellowish brown lactifers. Trama of pileus and stipe with abundant rosettes. Clamp connections absent.
Habitat and distribution: in forests dominated by fagaceous trees, 2-3 individuals growing together. July to September. Only known from Hunan.
Additional specimens examined: CHINA. Hunan: Sangzhi County, Badagong Mountain Nature Reserve, in forest dominated with Fagus lucida and Betula insignis, 8 Sept. 2020, Z.H. Chen, MHHNU 32523, MHHNU 32529 (both in MHHNU).
Note: the diagnostic features of this new species are the grayish green viscid pileus, the ellipsoid spores with zebroid ornamentation, and the pileipellis an ixolattice with olivaceous incrustations. It belongs to the subsect. Pyrogalini (Heilmann-Clausen et al. 1998) and is similar to L. blennius (Heilmann- Clausen et al. 1998) and L. fluens (Heilmann- Clausen et al. 1998) in the grayish green viscid pileus and spores with zebra-like ornamentation. Differing from the two related species, L. hunanensis has much slender basidiocarps and its basidiospores have typical zebroid ornamentation. Lactarius cinereus is another closely related species. It also has slender basidiocarps and grayish olivaceous viscid pileus, but its spores are much longer (Q=1.41-1.42) and have less zebroid ornamentation (Montoya et al. 2010). The spores of this new species are similar to those of L. circellatus, L. incrustatus and L. parallelus, but the latter three have stouter basidiocarps with less sticky pilei (Heilmann- Clausen et al. 1998; Lee et al. 2019; Wang 2019). Japanese L. glutininitens and L. avellaneus might be close due to the dull-colored viscid pileus and incrustations in the pileipellis (Imai 1935; Takahashi 2001; Wang & Liu 2010; observation on the holotype of L. glutininitens). These two species have much larger and more reticulate spores and lack greenish tinges (Wang & Liu 2010; observation on the holotype of L. glutininitens). This species has a strong temperate affinity. Its mycorrhizal partner is unknown, but the association with the trees of Betula could not be excluded. Considering all the other species in this species complex are not edible, we do not recommend to eat this species.
Lactarius polycystis Fang Wu, X.H. Wang & Z.H. Chen sp. nov.Fig. 8
Fig. 8

Fig. 8
Lactarius polycystis (MHHNU 30481, holotype).
A: Basidiocarps; B: Basidiospores; C: Pleuromacrocystidia; D: Cheilomacrocystidia; E: Pileipellis.
MycoBank MB 842238
DNA barcode ITS sequences: MK167402 (holotype), OL770163, OL770164.
Holotype: CHINA. Hunan: Shimen County, Huping Mountain Nature Reserve, Shenjingdong 15 Sept. 2009, Z.H. Chen, MHHNU 30481 (MHHNU!).
Etymology: the abundant typical macrocystidia.
Basidiocarps small to medium-sized. Pileus 18-40 mm diam., convex when young, becoming plano-convex with a depressed center, margin crenate; surface velvety, radially rugose, grayish cream, ochraceous brown to brick-brown, sometimes with an olivaceous tinge, often with paler or whitish patches. Lamellae adnate to decurrent, moderately crowded, 2-4 mm broad, pale yellow to light yellow. Stipe 12-40×3-10 mm, more or less cylindrical, central; surface dry, smooth, whitish to whitish cream. Context 2-3 mm thick, white. Taste and smell unknown. Latex white, rapidly turning yellow, finally staining lamellae brown.
Basidiospores (7.5) 8.0-8.6-10.0×(6.5) 7.0-8.0-9.0 µm [Q=(1.00) 1.02-1.08-1.18 (1.20)] (60/3/2), globose, subglobose, rarely broadly ellipsoid; ornamentation up to 3.5 μm high, mostly 1.5-2.8 μm high, of acute and heavy ridges and irregular warts, ridges branched and connected by wings, but very rarely forming closed meshes, often less amyloid at the top rendering a split appearance; plage distally amyloid. Basidia 45-62×10-16 µm, subclavate to clavate, 4-spored; sterigmata 4-7 µm long. Pleuromacrocystidia 30-113×8-15 µm, common, very variable in shape and size, smaller ones originating from the same depth as basidia, larger ones from subhymenium, and trama, subclavate, subfusiform, apex blunt, some constricted, with crystalline contents. Lamellar edge fertile; marginal cells 10-17×5-7 µm, clavate, hyaline; cheilomacrocystidia 25-35×6-10 µm, numerous, subfusiform, ventricose, with pale yellowish contents. Hymenophoral pseudocystidia scattered to common, 3-6 μm broad, cylindrical, tortuous, apex slightly enlarged or branching, hyaline. Pileipellis a trichoepithelium, 70-120 μm thick, with yellowish brown pigmentation; hyphae gelatinized, cylindrical or narrowly clavate in suprapellis, 25-54×3-7 μm, those in subpellis hyphoid or isodiametric, 10-30×6-18 μm. Stipitipellis a cutis to trichoderm, 30-50 μm thick; hyphae hyaline or pale yellowish brown, not gelatinized, terminal cells cylindrical or fusiform, 26-65×3-6 μm. Hymenophoral trama with abundant and conspicuous bright yellow to yellowish brown lactifers. Trama of pileus and stipe with abundant rosettes. Clamp connections absent.
Habitat and distribution: in fagaceous forests. Solitary or two individuals growing together. August to September. Central (Henan) and central-south China (Hunan).
Additional specimens examined: CHINA. Henan: Luanchuan County, Laojunshan, Zhaigou, Tenglongyu, in a fagaceous forest, alt. 1 250 m, 13 Aug. 2015, X.H. Wang, 3654 (KUN-HKAS 89927).
Note: Lactarius polycystis is unique for the presence of typical macrocystidia which is unusual in subg. Plinthogalus. The new species is a sibling of L. crassiusculus from Thailand (Le et al. 2007). Compared to the latter, L. polycystis has less crowded lamellae and larger spores with more prominent and less reticulate ornamentation. The latex of L. polycystis is yellow when bruised and not turning orange or pink as in L. crassiusculus. Lactarius crassiusculus reported by Chen & Zhang (2019) is a misidentification of the species. This is the second species with yellowing latex of the subgenus in China, after the newly published L. mirus (Wang et al. 2021). This species is not edible.
3 DISCUSSION
Except for L. austroscrobiculatus, all the five new species reported in this study are closely related with and morphologically similar to some known species. Nevertheless using multiple collections and DNA barcode gaps, as well as morphological characters, these species can be clearly distinguished. The finding of these species particularly enriches the species diversity of ectomycorrhizal fungi in central and eastern parts of China. This finding coincides with the prediction of Yang (2020): “Trends in fungal systematics: more and more taxa”.
Southern China harbors a large number of new and/or endemic fungal species (Wu et al. 2016; Cui et al. 2018; Dai et al. 2021; Zhang et al. 2021). Among the six species reported in this study, only L. hunanensis has strong affinity with temperate species. All the others, either have direction connection with tropical species, i.e. L. austroscrobiculatus, or have very close relatives in the tropics, e.g. L. cheilocystidiatus and L. polycystis. Together with other species found in southern China, e.g. Amanita mira, Cantharellus cerinoalbus, L. austrorostratus, L. formosus, Phlobopus portentosus and Russula purpureogracilis (Yang 2005; Song et al. 2017; Rossi et al. 2020; Zhao & Ji 2021), the species documented in this study will eventually help to establish a link between China and the other tropical regions in Asia and even Australasia.
Acknowledgments
We thank S.Q. Cao, Kunming Institute of Botany, Chinese Academy of Sciences for doing lab work. Prof. ZENG Niankai, Hainan Medical University, and Dr. ZHAO Kuan, Jiangxi Science and Technology Normal University, provided important specimens.
参考文献
Studies on the species of Lactarius from north Guangdong of China
Atlas of macrofungi in Hunan
The family Amanitaceae: molecular phylogeny, higher-rank taxonomy and the species in China
DOI:10.1007/s13225-018-0405-9 URL [本文引用: 1]
Diversity and systematics of the important macrofungi in Chinese forests
Phase determination from direct sequencing of length-variable DNA regions
DOI:10.1111/j.1471-8286.2006.01355.x URL [本文引用: 1]
BioEdit:a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT
Lactarius guangdongensis sp. nov. (Russulaceae, Russulales), a species of Lactarius sect. Deliciosi growing with a vulnerable five-needle pine, Pinus kwangtungensis
DOI:10.11646/phytotaxa.393.3.4 URL [本文引用: 1]
The genus Lactarius: Fungi of northern Europe. Vol. 2
North American species of Lactarius
Studies on the Agaricaceae of Japan. Ⅱ. Lactarius in Hokkaido
DOI:10.15281/jplantres1887.49.603 URL [本文引用: 1]
Recent developments in the MAFFT multiple sequence alignment program
DOI:10.1093/bib/bbn013 URL [本文引用: 1]
MEGA X: molecular evolutionary genetics analysis across computing platforms
DOI:10.1093/molbev/msy096 URL [本文引用: 1]
Lactarius in northern Thailand: 2. Lactarius subgen. Plinthogali
Taxonomic revision of the genus Lactarius (Russulales, Basidiomycota) in Korea
DOI:10.1007/s13225-019-00425-6 URL [本文引用: 3]
Mushroom poisoning outbreaks — China, 2020
DOI:10.46234/ccdcw2021.014 URL [本文引用: 1]
Mushroom poisoning outbreaks — China, 2021
Two Lactarius species associated with a relict Fagus grandifolia var. mexicana population in a Mexican montane cloud forest
Ectomycorrhizal (EM) fleshy fungi are being monitored in a population of Fagus grandifolia var. mexicana persisting in a montane cloud forest refuge on a volcano in a subtropical region of central Veracruz (eastern Mexico). The population of Fagus studied represents one of the 10 recognized forest fragments still housing this tree genus in Mexico. This is the first attempt to document EM fungi associated with this tree species in Mexico. We present evidence of the ectomycorrhizal symbiosis for Lactarius badiopallescens and L. cinereus with this endemic tree. Species identification of Lactarius on Fagus grandifolia var. mexicana was based on the comparison of DNAsequences (ITS rDNA) of spatiotemporally co-occurring basidiomes and EM root tips. The host of the EM tips was identified by comparison of the large subunit of the ribulose-bisphosphate carboxylase gene (rbcL). The occurrence of Lactarius badiopallescens and L. cinereus populations in the area of study represent the southernmost record known to date of these two species in North America and are new for the Neotropical Lactarius mycota. Descriptions coupled with illustrations of macro- and micromorphological features of basidiomes as well as photographs of ectomycorrhizas are presented.
Ectomycorrhizal fungal communities in endangered Pinus amamiana forests
Revisiting the host effect on ectomycorrhizal fungal communities: implications from host-fungal associations in relict Pseudotsuga japonica forests
DOI:10.1007/s00572-013-0504-0
PMID:23702643
[本文引用: 1]

Host identity is among the most important factors in structuring ectomycorrhizal (ECM) fungal communities. Both host-fungal coevolution and host shifts can account for the observed host effect, but their relative significance in ECM fungal communities is not well understood. To investigate these two host-related mechanisms, we used relict forests of Pseudotsuga japonica, which is an endangered endemic species in Japan. As with other Asian Pseudotsuga species, P. japonica has been isolated from North American Pseudotsuga spp. since the Oligocene and has evolved independently as a warm-temperate species. We collected 100 soil samples from four major localities in which P. japonica was mixed with other conifers and broadleaf trees. ECM tips in the soil samples were subjected to molecular analyses to identify both ECM fungi and host species. While 136 ECM fungal species were identified in total, their communities were significantly different between host groups, confirming the existence of the host effect on ECM fungal communities. None of the 68 ECM fungal species found on P. japonica belonged to Pseudotsuga-specific lineages (e.g., Rhizopogon and Suillus subgroups) that are common in North America. Most of ECM fungi on P. japonica were shared with other host fungi or phylogenetically close to known ECM fungi on other hosts in Asia. These results suggest that after migrating, Pseudotsuga-specific fungal lineages may have become extinct in small isolated populations in Japan. Instead, most of the ECM fungal symbionts on P. japonica likely originated from host shifts in the region.
Fungal biodiversity profiles 91-100
DOI:10.5252/cryptogamie-mycologie2020v41a4 URL [本文引用: 1]
Three new species of Lactarius (Russulaceae, Russulales) from northeastern China
DOI:10.1016/j.myc.2017.11.001 URL [本文引用: 1]
Cantharellus cerinoalbus - an edible fungus newly recorded in China
Lactarius subgenus Plinthogalus of Malaysia
Notes on new Agaricales of Japan 3
DOI:10.1007/BF02461218 URL [本文引用: 1]
Lactarius pleuromacrocystidiatus (Russulaceae), a novel species from India
DOI:10.1007/s12225-018-9788-8 URL [本文引用: 1]
Lactarius subg. Lactarius (Russulaceae) in Indian Himalaya: two new species with morphology and phylogenetic inferences
DOI:10.7872/crym/v39.iss4.2018.467 URL [本文引用: 2]
Agaricales of Indonesia. 3. New records of the genus Lactarius (Basidiomycota, Russulales) from Java
Type studies of Lactarius species published from China
DOI:10.1080/15572536.2007.11832585 URL [本文引用: 1]
Seven new species of Lactarius subg. Lactarius (Russulaceae) from southwestern China
Fungal biodiversity profiles 71-80
DOI:10.7872/crym/v39.iss4.2018.419 URL [本文引用: 2]
Two new species of Lactarius from Changbai and Lesser Khingan Mountains
Type study on Japanese species of Lactarius described by S
Revisiting the morphology and phylogeny of Lactifluus with three new lineages from southern China
DOI:10.3852/13-393 URL [本文引用: 2]
Lactarius vividus sp. nov. (Russulaceae, Russulales), a widely distributed edible mushroom in central and southern China
DOI:10.11646/phytotaxa.231.1.6 URL [本文引用: 1]
Phylogeny, biogeography and taxonomic re-assessment of Multifurca (Russulaceae, Russulales) using three-locus data
Lactifluus parvigerardii sp nov., a new link towards the pleurotoid habit in Lactifluus subgen. gerardii (Russulaceae, Russulales)
DOI:10.7872/crym.v33.iss2.2012.181 URL [本文引用: 1]
Russula griseocarnosa sp. nov. (Russulaceae, Russulales), a commercially important edible mushroom in tropical China: mycorrhiza, phylogenetic position, and taxonomy
DOI:10.1127/0029-5035/2009/0088-0269 URL [本文引用: 1]
Lactarius mirus sp. nov. (Russulaceae, Russulales), the first species of Lactarius subg. Plinthogalus with yellowing latex in China
Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics
Lactarius subgenus Russularia (Russulaceae) in South-East Asia: 3. new diversity in Thailand and Vietnam
DOI:10.11646/phytotaxa.207.3.1 URL [本文引用: 1]
The regionalization of Chinese flora
One hundred noteworthy boletes from China
DOI:10.1007/s13225-016-0375-8 URL [本文引用: 1]
Trends in fungal systematics: more and more taxa
Studies on the genus Lactarius S.F. Gray from China Ⅰ. New taxa of Lactarius
Multigene phylogeny and morphology reveal unexpectedly high number of new species of Cantharellus subgenus Parvocantharellus (Hydnaceae, Cantharellales) in China
DOI:10.3390/jof7020091 URL [本文引用: 1]
Atlas of macrofungi from Jingning of Zhejiang Province



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}