



广东南岭地区地处广东省北部,南岭山脉南端,地理位置位于23°53′-25°53′N,112°53′- 114°45′E,东西约192 km,南北约178 km,总面积为18 413.6 km2 (邵上等 2020)。南岭是重要的地域分界线和气候分界线,处于国家生态安全战略格局中的南方丘陵山地带的核心区域,属亚热带季风气候并兼具山地气候特色,降水充沛。南岭也是广东生物多样性最丰富的地区,保存有较好的亚热带季风常绿阔叶林、沟谷雨林等森林植被,是广东现今唯一的原始森林,森林覆盖率达73.8%,仅记录在南岭国家公园的高等植物就有324科1 522属5 105种(黄金玲等 2020;缪绅裕等 2020;邵上等 2020)。南岭生态保护核心区域具有原始生态风貌,是古热带动植物的避难所和近代东亚温带、亚热带植物的发源地(吴雪琴等 2013;宋斌等 2018)。受粤港澳大湾区辐射的带动力,成为大湾区的重要靠山,也是探索生态系统保护和资源合理利用平衡模式的重要区域(王发兴 2009;邵上等 2020;庄长伟等 2021)。南岭作为华南地区重要的森林生态区,是研究我国大型真菌物种多样性不可忽略的地区,近年我国对南岭地区的大型木材腐朽真菌进行了系统研究,发现了诸多新种和中国新记录种(吴芳等2020;戴玉成等2021)。目前南岭记录的大型真菌物种超1 275种,其中有52种大型真菌的模式产地是南岭,如:美丽金牛肝菌Aureoboletus formosus Ming Zhang & T.H. Li、栗褐金牛肝菌Aureoboletus marroninus T.H. Li & Ming Zhang、华南干巴菌Thelephora austrosinensis T.H. Li & T. Li、华南鸡油菌Cantharellus austrosinensis Ming Zhang, C.Q. Wang & T.H. Li和灰孔热带孔菌Tropicoporus ravidus Y.C. Dai & F. Wu 等(Zhang et al. 2015, 2019;宋斌等 2018;Li et al. 2019;Wu et al. 2022)。
牛肝菌隶属于担子菌门Basidiomycota、蘑菇纲Agaricomycetes、牛肝菌目Boletales,是大型担子菌中的一个重要类群,具有十分重要的生态、科研与经济价值。在生态上,绝大部分牛肝菌物种为外生菌根菌,可与植物形成共生关系,为植物提供营养成分,参与生态碳-氮循环,维护生态平衡(何青等 2021;黄兰兰等 2022);在科研上,由于牛肝菌形态特征多样,加上趋同进化、平行进化等各种复杂因素,使得早期基于形态及部分生理生化特征的牛肝菌分类系统,已难以真实反映其各类群的进化历史和亲缘关系,研究牛肝菌的分类和各类群间的进化关系成为了牛肝菌系统分类学研究中的热点和难点话题,受到广大科研工作者的关注(杨祝良 2013);牛肝菌中还有一些著名野生食用菌,如深受欧洲人喜爱的美味牛肝菌Boletus edulis Bull.,云南人钟爱的见手青——兰茂牛肝菌Lanmaoa asiatica G. Wu & Zhu L. Yang、玫红黄肉牛肝菌Butyriboletus roseoflavus (Hai B. Li & Hai L. Wei) D. Arora & J.L. Frank和目前已实现人工栽培的暗褐脉柄牛肝菌Phlebopus portentosus (Berk. & Broome) Boedijn等。牛肝菌中还含有多种生物活性物质,如多糖、萜类化合物、黄酮及多酚等,具有重要的经济和营养价值(李泰辉和宋斌 2002;Wu et al. 2019)。
本研究对南岭地区牛肝菌物种多样性进行了系统研究,对已报道的牛肝菌种类进行了整理统计和标本复查,对存疑或错误鉴定物种进行修订与注释,并基于nrLSU序列数据构建了南岭地区分布的牛肝菌物种系统发育树,明确了各物种的系统学位置,为物种的准确鉴定提供了分子证据。本研究还发现牛肝菌新种1个,并进行了描述与报道。
1 材料与方法
1.1 研究材料
1.2 形态学鉴定
野外拍摄标本生态照片,之后采集,并详细记录标本的生境信息和宏观形态特征,然后在40-50 ℃低温烘干处理。干标本作为显微特征观察材料,以5% KOH溶液作为浮载剂,在光学显微镜下观察和记录显微结构特征,随机选取测量20个成熟孢子的长和宽,以a-b × c-d表示,其中a、b代表孢子长度的最小值和最大值,c、d代表孢子宽度的最小值和最大值,Q值为长宽比,Qm为孢子长宽比的平均数和标准差。其他微观结构,如担子、囊状体、菌盖皮层及其末端细胞等均进行测量和记录。
1.3 分子系统学鉴定
对标本采用真菌基因组DNA快速抽提试剂盒(广州美基生物科技有限公司)提取总DNA,使用ITS1/ITS4 (White et al. 1990)和LR0R/LR5 (Vilgalys & Hester 1990)引物对,进行目的基因片段nrITS、nrLSU序列的PCR扩增。反应体系为25 μL,其中PCR mix 12 μL,10 mol/L引物各1 μL,DNA模板1 μL,用ddH2O定容至25 μL。热循环参数:94 ℃预变性5 min;94 ℃变性1 min,56 ℃ (ITS)/52 ℃ (LSU)退火1 min,72 ℃延伸90 s,35个循环;72 ℃延伸10 min,存于4 ℃。PCR反应产物送往北京六合华大基因科技有限公司广州分公司进行测序,利用Geneious软件进行序列拼接,在NCBI上进行序列比对,通过比对结果及文献查阅,选择和下载相关序列构建系统发育树。利用MAFFT在线比对网站进行多序列比对(Katoh et al. 2019),利用Bioedit软件进行序列的手动调整(Hall 1991),利用PhyloSuite_v1.2.2的ModelFinder选择序列矩阵的最佳模型,选择IQTREE进行最大似然法(maximum likelihood, ML)和Mrbayes进行贝叶斯法(Bayesian Inference, BI)构建系统发育树(Zhang et al. 2020)。
1.4 标本复查及信息分析
2 结果与分析
2.1 系统发育分析
基于南岭地区分布的60个物种标本、79条nrLSU序列,结合GenBank数据库下载的牛肝菌相关序列113条,构建了nrLSU序列矩阵,包含953个核酸位点,采用最大似然法和贝叶斯法构建系统发育树,以Scleroderma dictyosporum Pat.为发育树的外类群。结果(图1)显示,比对ML树与BI树,有相同的拓扑结构,结果仅展示ML树,将BI树的后验概率标记在节点的分支处。系统树包含牛肝菌科Boletaceae、圆孔牛肝菌科Gyroporaceae和乳牛肝菌科Suillaceae 3科,各科均为单系,且有较高的支持率。牛肝菌科包含25属53种,圆孔牛肝菌科包含1属4种,乳牛肝菌科包含1属3种。本研究所选样本在系统树上均得到较好区分,属间也形成独立主干分支,并获得较高支持率。在系统树上,牛肝菌科形成了7个独立的主干分支,故参考近年提出的牛肝菌科7亚科的分类系统(Wu et al. 2014, 2016;Li & Yang 2021)。
图1

图1
基于nrLSU序列构建的南岭牛肝菌最大似然树
图中已分别标示自展值(BS)和贝叶斯后验概率(BPP),自展值(BS)>50% (左)、贝叶斯后验概率(BPP)>0.90 (右)的数值标注于分支节点上. 在牛肝菌科中,不同颜色表示不同亚科
Fig. 1
The RAxML tree based on the nrLSU dataset of boletes in Nanling.
Posterior probabilities maximum likelihood support values BS values > 50% (left) and BPP values > 0.90 (right) are showed. Different colored clades within Boletaceae indicated different subfamilies.
2.2 物种复查及校对
通过对南岭地区牛肝菌资源的野外调查、馆藏标本复查与校对和名录汇编,确定南岭地区牛肝菌物种79种,隶属于3科31属,包含1新种;排除错误鉴定物种20种,排除无效名称物种8种;尚存在疑问物种25种。对排除和存疑物种进行了注释与分析,详细信息如下。
2.2.1 南岭地区已确定牛肝菌物种名录
*1. 粗糙擬粉孢牛肝菌Abtylopilus scabrosus Yan C. Li & Zhu L. Yang,凭证标本:GDGM72208。
*2. 重孔金牛肝菌Aureoboletus duplicatoporus (M. Zang) G. Wu & Zhu L. Yang,凭证标本:GDGM49451、GDGM71724。
#3. 美丽金牛肝菌Au. formosus Ming Zhang & T.H. Li,凭证标本:GDGM44441。
#4. 灰褐金牛肝菌Au. griseorufescens Ming Zhang & T.H. Li,凭证标本:GDGM28490。
5. 长柄金牛肝菌Au. longicollis (Ces.) N.K. Zeng & Ming Zhang,凭证标本:GDGM44734、GDGM49735。
#6. 栗褐金牛肝菌Au. marroninus T.H. Li & Ming Zhang,凭证标本:GDGM43288。
7. 小橙黄金牛肝菌Au. miniatoaurantiacus (Bi & Loh) Ming Zhang, N.K. Zeng & T.H. Li,凭证标本:GDGM43282。
*8. 肾孢金牛肝菌Au. nephrosporus G. Wu & Zhu L. Yang,凭证标本:GDGM76970。
#9. 萝卜味金牛肝菌Au. raphanaceus Ming Zhang & T.H. Li,凭证标本:GDGM75476。
10. 东方褐盖金牛肝菌Au. sinobadius Ming Zhang & T.H. Li,凭证标本:GDGM49670、GDGM49482、GDGM75477。
#11. 独生金牛肝菌Au. solus Ming Zhang & T.H. Li,凭证标本:GDGM70342。
*12. 纤细金牛肝菌Au. tenuis T.H. Li & Ming Zhang,凭证标本:GDGM77432。
13. 云南金牛肝菌Au. yunnanensis G. Wu & Zhu L. Yang,凭证标本:GDGM26359。
*14. 臧氏金牛肝菌Au. zangii X.F. Shi & P.G. Liu,凭证标本:GDGM75881。
*15. 梭孢南方牛肝菌Austroboletus fusisporus (Kawam. ex Imazeki & Hongo) Wolfe,凭证标本:GDGM77654。
16. 淡绿南方牛肝菌Aus. subvirens (Hongo) Wolfe,凭证标本:GDGM 27348。
*17. 黄肉条孢牛肝菌Boletellus aurocontextus Hirot. Sato,凭证标本:GDGM8842、GDGM8866、GDGM8891。
*18. 金色条孢牛肝菌B. chrysenteroides (Snell) Snell,凭证标本:GDGM72442。
19. 木生条孢牛肝菌B. emodensis (Berk.) Singer,凭证标本:GDGM49419、GDGM55155。
*20. 隐纹条孢牛肝菌B. indistinctus G. Wu, Fang Li & Zhu L. Yang,凭证标本:GDGM81359。
21. 深红条孢牛肝菌B. obscurococcineus (Höhn.) Singer,凭证标本:GDGM44784、GDGM 44761。
#22. 栗褐色牛肝菌Boletus castaneobrunneus Z.S. Bi,凭证标本:GDGM7969。
23. 灰盖牛肝菌Bo. griseiceps B. Feng, Y.Y. Cui, J.P. Xu & Zhu L. Yang,凭证标本:GDGM7527。
*24. 东方白牛肝菌Bo. orientialbus N.K. Zeng & Zhu L. Yang,凭证标本:GDGM7801。
25. 亚黄褐牛肝菌Bo. subfulvus Z.S. Bi,凭证标本:GDGM8120、GDGM8132。
26. 近浅灰色牛肝菌Bo. subgriseus Z.S. Bi,凭证标本:GDGM8122。
27. 紫牛肝菌Bo. violaceofuscus W.F. Chiu,凭证标本:GDGM8048、GDGM8819。
*28. 海南黄肉牛肝菌Butyriboletus hainanensis N.K. Zeng, Zhi Q. Liang & S. Jiang,凭证标本:GDGM75302。
*29. 玫红黄肉牛肝菌Bu. roseoflavus (Hai B. Li & Hai L. Wei) D. Arora & J.L. Frank,凭证标本:GDGM28480、GDGM29270。
*30. 象头山美牛肝菌Caloboletus xiangtoushanensis Ming Zhang, T.H. Li & X.J. Zhong,凭证标本:GDGM74885、GDGM75682。
#31. 辐射辣牛肝菌Chalciporus radiatus Ming Zhang & T.H. Li,凭证标本:GDGM 43305。
32. 柠檬黄辣牛肝菌Cha. citrinoaurantius Ming Zhang & T.H. Li,凭证标本:GDGM 44776。
*33. 窄囊裘氏牛肝菌Chiua angusticystidiata Y.C. Li & Zhu L. Yang,凭证标本:GDGM45987。
*34. 绿盖裘氏牛肝菌Chi. virens (W.F. Chiu) Y.C. Li & Zhu L. Yang,凭证标本:GDGM44758。
*35. 金红橙牛肝菌Crocinoboletus rufoaureus (Massee) N.K. Zeng, Zhu L. Yang & G. Wu,凭证标本:GDGM25887。
*36. 红褶牛肝菌Erythrophylloporus cinnabarinus Ming Zhang & T.H. Li,凭证标本:GDGM70536。
#37. 白黄圆孔牛肝菌Gyroporus alboluteus Ming Zhang & T.H. Li,凭证标本:GDGM25474。
*38. 蓝黑圆孔牛肝菌G. atrocyanescens Ming Zhang & T.H. Li,凭证标本:GDGM7781、GDGM7829、GDGM7992、GDGM75894、GDGM76540。
*39. 长囊体圆孔牛肝菌G. longicystidiatus Nagas. & Hongo,凭证标本:GDGM25857。
*40. 褐圆孔牛肝菌G. memnonius N.K. Zeng, H.J. Xie & M.S. Su,凭证标本:GDGM44779。
*41. 锥盖网孢牛肝菌Heimioporus conicus N.K. Zeng & Zhu L. Yang,凭证标本:GDGM77226。
42. 日本网孢牛肝菌H. japonicus (Hongo) E. Horak,凭证标本:GDGM43532、GDGM43542。
43. 中华网柄牛肝菌H. sinensis Ming Zhang, T.H. Li & X.N. Chen,凭证标本:GDGM44447。
*44. 皱半疣柄牛肝菌Hemileccinum rugosum G. Wu & Zhu L. Yang,凭证标本:GDGM27345、GDGM29053。
*45. 黑斑厚瓤牛肝菌Hourangia nigropunctata (W.F. Chiu) Xue T. Zhu & Zhu L. Yang,凭证标本:GDGM81356。
46. 兰茂牛肝菌Lanmaoa asiatica G. Wu & Zhu L. Yang,凭证标本:GDGM74901。
*47. 大盖兰茂牛肝菌L. macrocarpa N.K. Zeng, H. Chai & S. Jiang,凭证标本:GDGM77661。
48. 青木氏小绒盖牛肝菌Parvixerocomus aokii (Hongo) G. Wu, N.K. Zeng & Zhu L. Yang,凭证标本:GDGM43270。
49. 美丽褶孔菌牛肝菌Phylloporus bellus (Massee) Corner,凭证标本:GDGM70284、GDGM70500。
*50. 潞西褶孔牛肝菌Ph. luxiensis M. Zang,凭证标本:GDGM49753。
*51. 黑紫红孢牛肝菌Porphyrellus nigropurpureus (Hongo) Y.C. Li & Zhu L. Yang,凭证标本:GDGM80642。
52. 拟南牛肝菌Pseudoaustroboletus valens (Corner) Y.C. Li & Zhu L. Yang,凭证标本:GDGM80587。
*53. 褐糙粉末牛肝菌Pulveroboletus brunneoscabrosus Har. Takah.,凭证标本:GDGM43281。
54. 疽黄粉末牛肝菌Pu. icterinus (Pat. & C.F. Baker) Watling,凭证标本:GDGM56758、GDGM80591。
*55. 褐网柄牛肝菌Retiboletus fuscus (Hongo) N.K. Zeng & Zhu L. Yang,凭证标本:GDGM75280。
56. 灰褐牛肝菌R. griseus (Frost) Manfr. Binder & Bresinsky,凭证标本:GDGM7740、GDGM7741。
57. 黑网柄牛肝菌R. nigerrimus (R. Heim) Manfr. Binder & Bresinsky,凭证标本:GDGM45922。
*58. 灰盖罗扬牛肝菌Royoungia grisea Y.C. Li & Zhu L. Yang,凭证标本:GDGM75293、GDGM72439。
59. 褐鳞松塔牛肝菌Strobilomyces brunneolepidotus Har. Takah. & Taneyama,凭证标本:GDGM44426。
*60. 疣柄松塔牛肝菌S. verruculosus Hirot. Sato,凭证标本:GDGM80395。
61. 半裸松塔牛肝菌S. seminudus Hongo,凭证标本:GDGM88062。
**#62. 红褐赭黄牛肝菌Suillellus rufobrunneus Ming Zhang, S.X. He & T.H. Li, sp. nov.,凭证标本:GDGM74900、GDGM74924。
63. 美洲乳牛肝菌Suillus americanus (Peck) Snell,凭证标本:GDGM8863。
64. 黏盖乳牛肝菌Sui. bovinus (L.) Roussel,凭证标本:GDGM76110。
65. 点柄乳牛肝菌Sui. granulatus (L.) Roussel,凭证标本:GDGM27709。
*66. 滑皮乳牛肝菌Sui. huapi N.K. Zeng, R. Xue & Zhi Q. Liang,凭证标本:GDGM74883、GDGM76327。
67. 广东乳牛肝菌Sui. kwangtungensis Rui Zhang, X.F. Shi, P.G. Liu & G.M. Muell.,凭证标本:GDGM14116。
68. 近虎皮乳牛肝菌Sui. phylopictus Rui Zhang, X.F. Shi, P.G. Liu & G.M. Muell,凭证标本:GDGM70287。
#69. 近网纹乳牛肝菌Sui. subreticulatus Z.S. Bi,凭证标本:GDGM8112。
*70. 超群紫盖牛肝菌Sutorius eximius (Peck) Halling, Nuhn & Osmundson,凭证标本:GDGM76971。
71. 近圆孢粉孢牛肝菌Tylopilus balloui (Peck) Singer,凭证标本:GDGM56779。
72. 褐红粉孢牛肝菌T. brunneirubens (Corner) Watling & E. Turnbull,凭证标本:GDGM75276、GDGM75888、GDGM76803。
73. 新苦粉孢牛肝菌T. neofelleus Hong,凭证标本:GDGM8852、GDGM7839。
*74. 大津粉孢牛肝菌T. otsuensis Hongo,凭证标本:GDGM49481、GDGM55152、GDGM75275。
75. 类铅紫粉袍牛肝菌T. plumbeoviolaceoides T.H. Li, B. Song & Y.H. Shen,凭证标本:GDGM43535。
76. 褐金孢牛肝菌Xanthoconium fusciceps N.K. Zeng, Zhi Q. Liang & S. Jiang,凭证标本:GDGM44308。
*77. 中国金孢牛肝菌X. sinense G. Wu, Y.Y. Cui & Zhu L. Yang,凭证标本:GDGM43473。
*78. 拟小绒盖牛肝菌Xerocomus microcarpoides (Corner) E. Horak,凭证标本:GDGM74888。
*79. 黄盖臧氏牛肝菌Zangia citrina Y.C. Li & Zhu L. Yang,凭证标本:GDGM75404、GDGM75680、GDGM 75897。
注:*表示南岭新记录种;**表示新种;#表示以南岭为模式产地的物种。
2.2.2 排除物种名录
1. 网翼南方牛肝菌Austroboletus dictyotus (Boed.) Wolf.,标本复查实为梭孢南方牛肝菌Aus. fusisporus (Kawam. ex Imazeki & Hongo) Wolfe,Aus. dictyotus在华南地区有分布(Wu et al. 2016),在南岭地区尚未被发现。
2. 新柔南方牛肝菌Aus. malaccensis (Pat. & C.F. Baker) Wolfe,标本复查实为梭孢南方牛肝菌Aus. fusisporus (Kawam. ex Imazeki & Hongo) Wolfe。
3. 厚鳞条孢牛肝菌Boletellus ananas (M.A. Curtis) Murrill,标本复查实为黄肉条孢牛肝菌B. aurocontextus Hirot. Sato。
4. 凤梨盖条孢牛肝菌B. ananiceps (Berk.) Sing,标本复查实为黄肉条孢牛肝菌B. aurocontextus Hirot. Sato。
5. 狭孢条孢牛肝菌B. stenosporus T.H. Li et Zhang,无效名称。
6. 双色牛肝菌Boletus bicolor Peck.,已修订为兰茂牛肝菌L. asiatica G. Wu & Zhu L. Yang。
7. 美味牛肝菌Bo. edulis Bull.,标本复查实为灰盖牛肝菌B. griseiceps B. Feng, Y.Y. Cui, J.P. Xu & Zhu L. Yang。
8. 苦牛肝菌Bo. felleus Bull.,已修订为新苦粉孢牛肝菌T. neofelleus Hong。
9. 浅灰色牛肝菌Bo. griseus Frost,已修订为褐网柄牛肝菌R. fuscus (Hongo) N.K. Zeng & Zhu L. Yang。
10. 皱盖牛肝菌Bo. hortonii A.H. Sm. & Thiers,标本复查实为皱半疣柄牛肝菌He. rugosum G. Wu & Zhu L. Yang。
11. 紫盖牛肝菌Bo. inedulis (Murrill) Murrill,标本复查实为海南黄肉牛肝菌Bu. hainanensis N.K. Zeng, Zhi Q. Liang & S. Jiang。
12. 小美牛肝菌Bo. speciosus Frost,已修订为玫红黄肉牛肝菌Bu. roseoflavus (Hai B. Li & Hai L. Wei) D. Arora & J.L. Frank。
13. 小孢褐小牛肝菌Fuscoboletinus microsporus T.H. Li,无效名称。
14. 松林褐小牛肝菌F. pinetorum (Chiu) T.H. Li,无效名称。
15. 中国褐小牛肝菌F. sinicus Li,无效名称。
16. 广东圆孔牛肝菌Gyrodon guangdongensis T.H. Li,无效名称。
17. 褐圆孔牛肝菌Gyroporus castaneus (Bull.) Quél.,对早期的标本复查显示南岭地区分布样本至少存在2个物种, G. longicystidiatus Nagas. & Hongo和G. memnonius N.K. Zeng, H.J. Xie & M.S. Su。
18. 蓝圆孔牛肝菌G. cyanescens (Bull.) Quél.,标本复查实为蓝黑圆孔牛肝菌G. atrocyanescens Ming Zhang & T.H. Li。
19. 拟白圆孔牛肝菌G. pseudolacteus G. Moreno, Carlavilla, Heykoop, Manjón & Vizzini,标本复查实为蓝黑圆孔牛肝菌G. atrocyanescens Ming Zhang & T.H. Li。
20. 白疣柄牛肝菌Leccinum albellum (Peck) Singer,标本复查结果显示该种可能是近年描述于中国福建地区的东方白牛肝菌B. orientialbus N.K. Zeng & Zhu L. Yang。
21. 红黄褶孔牛肝菌Phylloporus rhodoxanthus (Schwein.) Bres.,已修订为美丽褶孔菌牛肝菌Ph. bellus (Massee) Corner。
22. 金粒粉末牛肝菌Pulveroboletus auriflammeus (Berk. & M.A. Curtis) Singer,已修订为小橙黄金牛肝菌Aur. miniatoaurantiacus (Bi & Loh) Ming Zhang, N.K. Zeng & T.H. Li。
23. 绒毛松塔牛肝菌Strobilomyces echinatus Beeli,复查后近似于S. brunneolepidotus Har. Takah. & Taneyama。
24. 南岭乳牛肝菌Suillus nanlingensis T.H. Li,无效名称。
25. 虎皮乳牛肝菌Sui. pictus (Peck) A.H. Sm. & Thiers,已修订为Sui. phylopictus R. Zhang, X.F. Shi, P.G. Liu & G.M. Mueller。
26. 褐金孢牛肝菌参照种Xanthoconium aff. affine (Peck) Singer,已修订为中国金孢牛肝菌 Xan. sinense G. Wu, Y.Y. Cui & Zhu L. Yang。
27. 小孢绒盖牛肝菌Xerocomus braohysporuo Li,无效名称。
28. 多褶绒盖牛肝菌Xer. satisfactus (Corn.) T.H. Li,无效名称。
2.2.3 存疑物种
1. 褐盖牛肝菌Boletus brunneissimus Chiu,缺少凭证标本信息。
2. 红脚牛肝菌Bo. erythropus (Fr.: Fr.) Krombh,缺少凭证标本信息,疑似此文描述的新物种。
3. 兄弟牛肝菌Bo. fraternus Peck,缺少凭证标本信息。
4. 褐变牛肝菌Bo. indecisus Peck,复查后形态上存在差异。
5. 褐黄牛肝菌Bo. luridus Schaeff.,复查结果显示形态描述上存在差异。
6. 血红牛肝菌Bo. rubellus Krombh.,缺少凭证标本信息,现更名为Hortiboletus rubellus (Krombh.) Simonini, Vizzini & Gelardi。
7. 红网柄牛肝菌Bo. satanas Lenz,缺少凭证标本信息。
8. 敏感牛肝菌Bo. sensibilis Peck.,缺少凭证标本信息。
9. 硬牛肝菌Bo. spraguei Frost,复查后形态上存在差异,现更名为Bo. hypohaematicus Singer。
10. 绒盖牛肝菌Bo. subtomentosus L.,该种主要在欧洲分布,在南岭地区是否真实存在仍需要研究。
11. 类虫形牛肝菌Bo. vermiculosoides A.H. Sm. & Thiers,复查后形态上存在差异。
12. 粘柄褐孔小牛肝菌Fuscoboletinus glandulosus (Peck) Pomerl. & A.H. Sm.,复查凭证标本在显微结果上存在差异。
13. 马来西亚圆孔牛肝菌Gyroporus malesicus Corner,复查凭证标本结果显示微观结构上存在差异。
14. 东方褶孔牛肝菌短孢变种Phylloporus orientalis var. brevisporus Corner,缺少凭证标本信息。
15. 硫黄褶孔牛肝菌Ph. sulphureus (Berk.) Singer.,缺少凭证标本信息。
16. 黄粉末牛肝菌Pulveroboletus ravenelii (Berk. & M.A. Curtis) Murrill,研究表明该种为一个复合种群,仍需进一步研究以澄清。
17. 混杂松塔牛肝菌Strobilomyces confusus Singer,复查后形态上存在差异。
18. 绒柄松塔牛肝菌S. floccopus (Vahl: Fr.) Karst.,复查后形态上存在差异。该物种常分布在欧洲、东亚和北美洲等地。
19. 乳流乳牛肝菌Sui. lactifluus (With.) A.H. Sm. & Thiers,缺少凭证标本信息。
20. 琥珀乳牛肝菌Sui. placidus (Bull.) Kuntze,复查后形态上存在差异,有研究表明分布在热带或亚热带的Sui. placidus (Bull.) Kuntze实为滑乳牛肝菌Sui. huapi N.K. Zeng, R. Xue & Zhi Q. Liang。
21. 绒毛乳牛肝菌Sui. tomentosus Singer, Snell & E.A. Dick,复查后形态上存在差异。
22. 栗色粉孢牛肝菌Tylopilus castaneiceps Hongo,缺少凭证标本信息。
23. 橄榄红粉孢牛肝菌T. olivaceirubens Corn.,缺少凭证标本信息,该物种与大津粉孢牛肝菌 T. otsuensis Hongo实为同物异名。
24. 白褐绒盖牛肝菌Xerocomus albobrunneus Heinem. & Gooss.-Font.,缺少凭证标本信息。
25. 红绒盖牛肝菌Xe. chrysenteron (Bull.) Quél.,缺少凭证标本信息。
2.3 新种描述
红褐赭黄牛肝菌 新种 图2
图2

图2
红褐赭黄牛肝菌
A,B:子实体及生境. C:担孢子. D:担子. E:侧身囊状体及管缘囊状体. 比例尺=10 μm
Fig. 2
Suillellus rufobrunneus.
A, B: Basidiomata and habitat. C: Basidiospores. D: Basidia. E: Pleurocystidia and cheilocystidia. Scale bars=10 μm.
Suillellus rufobrunneus Ming Zhang, S.X. He & T.H. Li, sp. nov. Fig. 2
Fungal Name FN571221
Etymology: The epithet “rufobrunneus” refers to the reddish brown colored pileus.
Holotype: CHINA, Guangdong Province, Shaoguan City, Renhua County, Danxiashan National Nature Reserve, 113°45′E, 25°03′N, alt. 240 m, 9 September 2018, Ming Zhang (GDGM74900, holotype; ITS-OP700054, 28S-OP700056).
Pileus 3-9 cm broad, subhemispheric at first, then becoming convex to plane, surface dry, dull red, greyish red, brownish red, reddish red to dark brown (8B4-8B6, 9D7-9D8, 9E7-9E8, 9F7-9F8), velutinous to floccos. Pileus context 5-20 mm thick in the center, fleshy, soft, pale yellow, yellowish white to white (2A1-2A3), turning blue (20A3-20A6, 24A3-24A6, 20A5-24A5) when exposed. Tubes 10-15 mm long, pastel yellow to yellow (1A1-2A8), staining blue when exposed; pores angular, 0.3-0.5 mm, pale yellow, pastel yellow to light yellow when young, changing blue when bruised. Stipe 2-7×1.5-3 cm, tapering upwards, central, solid, red to dark purplish red (10C8-10E8, 12C5-12C7), pale yellow to yellow at apex, with longitudinal and slight reticula all over the stipe; staining blue when injured; basal mycelium white. Odor indistinct. Taste mild.
Basidiospores 11-15×5.5-7 μm, [Q=1.73-2.32, Qm=2.05±0.15], ellipsoid and subfusiform in side view, inequilateral, smooth, yellowish brown in 5% KOH, inamyloid. Basidia 30-46×11-15 μm, clavate, sinuous, 4-sterigmate, sterigmata up to 4.3 µm long. Hymenophoral trama bilaterality. Pleurocystidia 34-53×8-10 µm, scattered, conspicuous, fusiform to clavate, thin-walled, smooth. Cheilocystidia similar to cheilocystidia in shape and size. Pileipellis an interwoven trichodermium to subcutis, composed of interwoven filamentous hyphae, 4-7 μm wide, yellowish to hyaline in 5% KOH; terminal cells 29-72×4-13 μm, cylindrical, clavate or nearly fusoid. Stipitipellis a tangled layer of repent to suberect hyphae, 3-7 μm wide, pale yellowish to hyaline in 5% KOH. Stipe trama composed of densely longitudinally arranged hyphae, 4-10 μm wide, pale yellowish to hyaline in 5% KOH. Clamp connections absent in all parts of basidiome.
Habitat, ecology and distribution: Solitary or scattered on the ground with humus and debris in broadleaf forests dominated by Fagaceae, alt. 500-800 m, currently only known from southern China.
Additional specimens examined: CHINA, Guangdong Province, Shaoguan City, Renhua County, Danxiashan National Nature Reserve, 113°45′E, 25°03′N, alt. 240 m, 9 September 2018, Ming Zhang (GDGM74924; ITS-OP700055, 28S-OP700057).
Notes: Suillellus rufobrunneus is characterized by its dry and brownish red to dark brown pileus, red to dark purplish red stipe with a pale yellow to yellow apex, with longitudinal and slight reticula over the stipe and bluing when injured. Morphologically, Su. subamygdalinus Kuan Zhao & Zhu L. Yang is similar to Su. rufobrunneus, and both of them have brownish red pileus, longitudinal reticula stipe, and all the basidiomata turning blue when bruised. However, Su. subamygdalinus differs in its longitudinal reticula, narrower basidia 34-48×8-11 μm, and larger cheilocystidia 38-56×7-10 μm (Wu et al. 2016).
菌盖直径3-9 cm,初期近半球形,后期凸镜形至平展;表面干燥,暗红色、红褐色、棕红色至深棕色,具绒毛。菌盖中心厚5-20 mm,肉质,柔软,淡黄色、黄白色至白色,伤变淡蓝色至蓝色。菌管长10-15 mm,淡黄色至黄色,伤后变蓝色;管口角形,0.3-0.5 mm,幼时淡黄色至鲜黄色,受伤后变蓝色。菌柄2-7×1.5-3 cm,向上逐渐变细,中生,实心,红色至深紫红色,顶端靠近菌管处黄色至淡黄色,表面有浅的纵向网纹,伤时变蓝,基部菌丝白色。气味不明显。味道温和。
担孢子11-15×5.5-7 μm,[Q=1.73-2.32,Qm=2.05±0.15],椭圆形,近纺锤形,不等边,光滑,在5% KOH中呈黄褐色,非淀粉质。担子30-46×11-15 μm,棒状,弯曲,4个担子小梗,担子小梗长达4.3 μm。子实层菌髓平行。侧生囊状体34-53×8-10 μm,明显,梭形或棍棒状,薄壁,光滑。管缘囊状体形状和大小与侧生囊状体相似。菌盖皮层近毛皮层状,菌丝轻微交织,薄壁,直径4-7 μm,在5% KOH中呈淡黄色至透明;菌盖皮层末端细胞29-72×4-13 μm,圆柱形、棍棒状或近纺锤形。菌柄皮层由匍匐至近直立菌丝组成,宽3-7 μm,淡黄色至透明,在5% KOH中呈淡黄色至透明。菌柄菌髓由密集的纵向排列菌丝组成,宽4-10 μm,在5% KOH中呈淡黄色至透明。锁状联合未见。
生境信息:单生或散在以壳斗科植物为主的阔叶林中地上,海拔500-800 m。
中国分布:广东南岭。
世界分布:中国。
红褐赭黄牛肝菌主要区别特征在于其红褐色至深棕色的菌盖表面,红色至深紫红色菌柄顶端浅黄色至黄色,表面具纵向浅网纹,受伤后变蓝(图2)。在形态上,红褐赭黄牛肝菌与Su. subamygdalinus较为相近,在系统发育关系上也与之相近,它们都具有红褐色的菌盖,菌柄表面都有纵向网纹,受伤时变蓝色。不同点在于Su. subamygdalinus菌柄上具有更长且更深的网纹,相对较窄的担子(34-48×8-11 μm)和相对较长的囊状体(38-56×7-10 μm) (Wu et al. 2016)。基于该新种与其近缘物种的nrITS序列,通过最大似然法和贝叶斯法构建系统发育树,比对ML树与BI树,有相同的拓扑结构,图3展示ML树,在系统发育树上,红褐赭黄牛肝菌形成独立的分支,并且具有高支持率。
图3
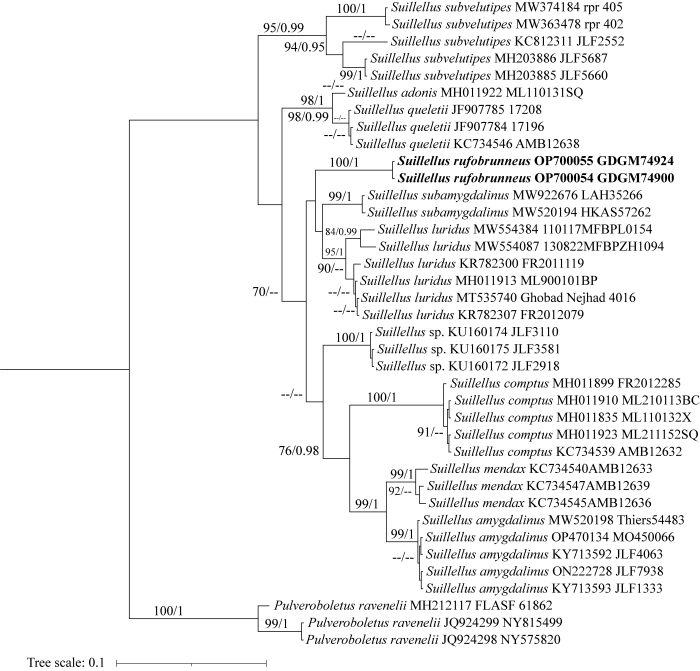
图3
基于nrITS序列的最大似然树
比对ML树与BI树,有相同的拓扑结构,仅展示ML树,图中已分别标示自展值(BS)和贝叶斯后验概率(BPP),自展值(BS)>70% (左)、贝叶斯后验概率(BPP)>0.90 (右)的数值标注于分支节点上
Fig. 3
The RAxML tree based on the nrITS dataset.
Posterior probabilities maximum likelihood support values. BS values > 70% (left) and BPP values > 0.90 (right) are showed. The new species are in bold.
3 结论与讨论
通过研究初步证明广东南岭地区分布牛肝菌3科(包括牛肝菌科Boletaceae、圆孔牛肝菌科Gyroporaceae和乳牛肝菌科Suillaceae),31属,79种,描述于中国本土的物种有55种,约占南岭牛肝菌物种数量的2/3;其中,以南岭为模式产地报道的物种有10种,包括本研究发现的1新种,南岭新记录种37种。对原有报道物种进行了复查与修订,排除了一些错误鉴定物种,对存疑物种进行了注释,并提供了大部分物种的分子证据,为准确认识南岭地区牛肝菌物种多样性提供基础数据。
牛肝菌是一个庞大的真菌类群,物种资源丰富,加之趋同进化、平行进化等因素存在,使得对牛肝菌的认识成为难题。早期由于研究的不足和对物种认识的局限性,使得一些形态相似的本土物种被错误地冠以欧美地区物种名称,如新苦粉孢牛肝菌T. neofelleus、褐网柄牛肝菌R. fuscus及玫红黄肉牛肝菌Bu. roseoflavus等分别被错误地鉴定为苦牛肝菌Bo. felleus、浅灰色牛肝菌Bo. griseus和小美牛肝菌Bo. speciosus;而一些物种的区分,需要借助分子序列才能完成,如皱盖牛肝菌Bo. hortonii与皱半疣柄牛肝菌He. rugosum;褐圆孔牛肝菌G. castaneus与G. memnonius;琥珀乳牛肝菌Sui. placidus与滑皮乳牛肝菌Sui. huapi等。同时,南岭地区牛肝菌还存在名称滥用或不合格发表等现象,如报道中记载的一些名称:小孢褐小牛肝菌Fuscoboletinus microsporus T.H. Li、南岭乳牛肝菌Suillus nanlingensis T.H. Li和小孢绒盖牛肝菌Xerocomus braohysporuo Li等(李泰辉等 2003),根据《国际藻类、菌物和植物命名法规》,这些物种名称均为不合格发表,因此予以排除。此外,在本研究中,还存在一些存疑物种,如:红网柄牛肝菌Bo. satanas、敏感牛肝菌Bo. sensibilis、红脚牛肝菌Bo. erythropus及褐变牛肝菌Bo. indecisus等,由于缺乏凭证标本或标本状况很差,这些物种是否在南岭地区有分布还存在疑问,需进一步广泛采集标本以求证。
牛肝菌绝大多数为外生菌根真菌,可与壳斗科Fagaceae、龙脑香科Dipterocarpaceae、豆科Leguminosae、杜鹃花科Ericaceae、桃金娘科Myrtaceae和松科Pinaceae等植物形成共生关系(Binder & Hibbett 2006),南岭地区丰富的牛肝菌资源与区域内具有丰富的宿主植物密切相关,南岭地区分布有以壳斗科植物甜槠Castanopsis eyrei、鹿角锥C. lamontii、罗浮锥C. faberi、米槠C. carlesii、青冈Cyclobalanopsis glauca等和松科植物广东松Pinus kwangtungensis和马尾松P. massoniana等为优势建群树种的典型植被类型,为牛肝菌的生长提供了有利条件;反之,丰富的牛肝菌资源在一定程度上可作为森林生态健康的指示凭证,侧面反映森林生态状态(杨汝荣 2000;徐卫等 2022;李挺等 2023)。
南岭牛肝菌资源是南岭生态屏障中生物多样性的重要组成部分,对南岭地区牛肝菌的科学复查,摸清南岭地区牛肝菌物种多样性情况,揭示南岭地区牛肝菌资源的丰富度和独特性,掌握南岭牛肝菌物种的分布与发生等动态变化规律,可为后续开展牛肝菌物种保护、探讨南岭森林生态功能和区域内物种协同进化研究提供数据基础。
参考文献
Macrofungus flora of Guangdong Province
Macrofungus flora of the mountainous district of North Guangdong
Molecular systematics and biological diversification of Boletales
Historical patterns of morphological evolution and ecology in the Boletales are largely unresolved but appear to involve extensive convergence. We studied phylogenetic relationships of Boletales based on two datasets. The nuc-lsu dataset is broadly sampled and includes roughly 30% of the described species of Boletales and 51 outgroup taxa across the Hymenomycetes. The multigene dataset (nuc-ssu, nuc-lsu, 5.8S, mt-lsu, atp6) sampled 42 key species of Boletales in a framework of 14 representative Hymenomycetes. The Boletales are strongly supported as monophyletic in our analyses on both datasets with parsimony, maximum likelihood and Bayesian approaches. Six major lineages of Boletales that currently are recognized on subordinal level, Boletineae, Paxillineae, Sclerodermatineae, Suillineae, Tapinellineae, Coniophorineae, received varied support. The backbone of the Boletales was moderately resolved in the analyses with the nuc-lsu dataset, but support was strong for most major groups. Nevertheless, most brown-rot producing forms were placed as a paraphyletic grade at the base of the Boletales. Analyses on the multigene dataset confirm sister group relationships among Boletales, Agaricales and Atheliales. Boletineae and Suillineae received the highest support values; Paxillineae and Sclerodermatineae were not consistently resolved as monophyletic groups. The Coniophorineae were not monophyletic in any analyses. The Tapinellineae consisting of morphologically diverse brown-rotting fungi forms the basal group in the Boletales. We performed ancestral state reconstruction with BayesMultiState, which suggested that the ancestor of the Boletales was a resupinate or polyporoid saprotrophic fungus, producing a brown-rot.
Diversity and systematics of the important macrofungi in Chinese forests
BioEdit:a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/ NT
Mitigation effect and regulation mechanism of ectomycorrhizal on plant under abiotic stress: a review
The core resources and scientific protection values of Guangdong Nanling National Park
Mycorrhizal synthesis of early stage fungi with native and exotic pines
Terms and scientific names of fungi
MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization
DOI:10.1093/bib/bbx068 URL [本文引用: 1]
Authors of fungal names. Index of Fungi Supplement
Methuen handbook of colour
Thelephora austrosinensis (Thelephoraceae), a new species close to T. ganbajun from southern China
DOI:10.11646/phytotaxa.471.3 URL [本文引用: 1]
Community structures and influencing factors of soil fungi associated with typical vegetations in Nanling Mountains
Species and distribution of edible porcini in China
Atlas of macrofungi in Chebaling
Fungal resources in Guangdong Nanling National Nature Reserve
In:
The boletes of China:Tylopilus s
Plant diversity and its conservation values of Guangdong Nanling National Park
Research and practice of ecological protection and restoration of mountain-river-forest-farmland-lake-grassland system in Nanling mountains area, northern Guangdong Province
The macrofungal diversity in the Nanling Mountains
DOI:10.13284/j.cnki.rddl.003053

The Nanling Mountain region of South China is very rich in macrofungal resources, and also is the largest oasis near 25 degrees north latitude. The macrofungal diversity in the Nanling Mountains was introduced in this paper. 37 species originally described from the type localities in the Nanling were listed, including the Latin name, Chinese name, habitat and distribution; some misidentification or questionable species were annotated; the dominant families or genera, floristic elements, resource protection and sustainable utilization in the Nanling Mountains were analyzed. Strengthening the researches and protection of macrofungal diversity in the Nanling Mountains is greatly important not only for scientific significance, but also for ecosystem stability, regional environmental safety and future industrial development.
Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species
DOI:10.1128/jb.172.8.4238-4246.1990
PMID:2376561
[本文引用: 1]
Detailed restriction analyses of many samples often require substantial amounts of time and effort for DNA extraction, restriction digests, Southern blotting, and hybridization. We describe a novel approach that uses the polymerase chain reaction (PCR) for rapid simplified restriction typing and mapping of DNA from many different isolates. DNA fragments up to 2 kilobase pairs in length were efficiently amplified from crude DNA samples of several pathogenic Cryptococcus species, including C. neoformans, C. albidus, C. laurentii, and C. uniguttulatus. Digestion and electrophoresis of the PCR products by using frequent-cutting restriction enzymes produced complex restriction phenotypes (fingerprints) that were often unique for each strain or species. We used the PCR to amplify and analyze restriction pattern variation within three major portions of the ribosomal DNA (rDNA) repeats from these fungi. Detailed mapping of many restriction sites within the rDNA locus was determined by fingerprint analysis of progressively larger PCR fragments sharing a common primer site at one end. As judged by PCR fingerprints, the rDNA of 19 C. neoformans isolates showed no variation for four restriction enzymes that we surveyed. Other Cryptococcus spp. showed varying levels of restriction pattern variation within their rDNAs and were shown to be genetically distinct from C. neoformans. The PCR primers used in this study have also been successfully applied for amplification of rDNAs from other pathogenic and nonpathogenic fungi, including Candida spp., and ought to have wide applicability for clinical detection and other studies.
Shaoguan area of Northern Guangdong: the strategic thinking of constructing the second regional economic center of Guangdong
Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR protocols, a guide to methods and applications
Polypore diversity in South China
Global diversity and systematics of Hymenochaetaceae with poroid hymenophore
DOI:10.1007/s13225-021-00496-4 [本文引用: 1]
Resource diversity of Chinese macrofungi: edible, medicinal and poisonous species
DOI:10.1007/s13225-019-00432-7 [本文引用: 1]
Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae
DOI:10.1007/s13225-014-0283-8 URL [本文引用: 1]
One hundred noteworthy boletes from China
DOI:10.1007/s13225-016-0375-8 URL [本文引用: 4]
Genetic diversity of natural and planted populations of Tsoongiodendron odorum from the Nanling Mountains
Distribution pattern of plant community species diversity along altitudinal gradients in Nanling Mountains, Guangdong
Biodiversity and ecosystem protection in Nanling Mountain region and environment safety
Fungal taxonomy in the genomics era: opportunities and challenges
PhyloSuite: an integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies
DOI:10.1111/1755-0998.13096
PMID:31599058
[本文引用: 1]
Multigene and genomic data sets have become commonplace in the field of phylogenetics, but many existing tools are not designed for such data sets, which often makes the analysis time-consuming and tedious. Here, we present PhyloSuite, a (cross-platform, open-source, stand-alone Python graphical user interface) user-friendly workflow desktop platform dedicated to streamlining molecular sequence data management and evolutionary phylogenetics studies. It uses a plugin-based system that integrates several phylogenetic and bioinformatic tools, thereby streamlining the entire procedure, from data acquisition to phylogenetic tree annotation (in combination with iTOL). It has the following features: (a) point-and-click and drag-and-drop graphical user interface; (b) a workplace to manage and organize molecular sequence data and results of analyses; (c) GenBank entry extraction and comparative statistics; and (d) a phylogenetic workflow with batch processing capability, comprising sequence alignment (mafft and macse), alignment optimization (trimAl, HmmCleaner and Gblocks), data set concatenation, best partitioning scheme and best evolutionary model selection (PartitionFinder and modelfinder), and phylogenetic inference (MrBayes and iq-tree). PhyloSuite is designed for both beginners and experienced researchers, allowing the former to quick-start their way into phylogenetic analysis, and the latter to conduct, store and manage their work in a streamlined way, and spend more time investigating scientific questions instead of wasting it on transferring files from one software program to another.© 2019 John Wiley & Sons Ltd.
A new species and a new combination of Caloboletus from China
DOI:10.11646/phytotaxa.309.2 URL [本文引用: 1]
Aureoboletus formosus, a new bolete species from Hunan Province of China
DOI:10.1007/s11557-015-1022-6 URL [本文引用: 2]
Phylogenetic overview of Aureoboletus (Boletaceae, Boletales), with descriptions of six new species from China
DOI:10.3897/mycokeys.61.47520
URL
[本文引用: 2]
In this study, species relationships of the genus Aureoboletus were studied, based on both morphological characteristics and a four-gene (nrLSU, tef1-a, rpb1 and rpb2) phylogenetic inference. Thirty-five species of the genus have been revealed worldwide, forming eight major clades in the phylogenetic tree, of which twenty-four species have been found in China, including six new species: A. glutinosus, A. griseorufescens, A. raphanaceus, A. sinobadius, A. solus, A. velutipes and a new combination A. miniatoaurantiacus (Bi & Loh) Ming Zhang, N.K. Zeng & T.H. Li proposed here. A key to 24 known Chinese species has been provided.
New insights into the genus Gyroporus (Gyroporaceae, Boletales), with establishment of four new sections and description of five new species from China
DOI:10.1080/21501203.2022.2094012
PMID:35938079
[本文引用: 1]
Species of from southern China were studied in this study. Based on morphology and molecular phylogenetic analyses of DNA sequences from the nuclear ribosomal internal transcribed spacer (ITS), the nuclear ribosomal large subunit (nrLSU), and the mitochondrial adenosine triphosphate ATP synthase subunit 6 ( was divided into four main branches in the phylogenetic tree, and four sections were firstly proposed i.e. sect.. sect.. sect. and. sect.. Five new species, i.e. and, were revealed from China, and their phylogenetic positions were also analysed. Among them, and were nested into the sect., although morphologically similar to and, with obvious cyanescent oxidation reactions, were nested into the sect. ; and characterised by its long cystidia and was nested into the sect.. The new species were formally described and illustrated in the present study, and a key to the sections and species of in China was provided.© 2022 The Author(s). Published by Informa UK Limited, trading as Taylor & Francis Group.
Planning and construction strategy in priority area of biodiversity conservation in Nanling Mountain of Guangdong
中国森林大型真菌重要类群多样性和系统学研究
广东南岭国家级自然保护区的真菌资源调查研究
南岭地区观光木自然和人工迁地保护种群的遗传多样性
DOI:10.3724/SP.J.1003.2013.09138
[本文引用: 1]
迁地保护是珍稀濒危植物保护的重要措施。观光木(Tsoongiodendron odorum)是古老的孑遗树种, 被列为国家二级重点保护植物, 营建迁地保护林是对其进行保护的重要手段, 但已经营建的迁地保护林每个种群目前只保存下来几十株个体。为了评价这些迁地保护林的遗传多样性现状, 作者采用ISSR分子标记对南岭地区观光木3个人工迁地保护种群和4个自然种群的遗传多样性进行了比较。结果显示: 16条ISSR引物共扩增出362个条带, 其中301个为多态条带, 多态条带百分比(P)为83.2%, 各种群P值为37.9-62.2%, 平均为53.1%, 表明观光木在物种和种群水平都具有较高的遗传多样性。自然种群总体P值和Shannon信息指数(I)(80.9%, 0.3629)均高于人工迁地保护种群(66.6%, 0.2990), 说明人工迁地保护种群遗传多样性较低。种群结构分析表明3个人工迁地保护种群均有可能来源于源口自然种群。4个自然种群间遗传分化系数(G<sub>ST</sub>)为0.2495, 表明自然种群间存在显著遗传隔离, 基因流受阻和生态环境差异是造成种群遗传分化的主要原因。在今后的迁地保护工作中, 我们建议从不同生态地理区收集种质材料, 并在不同生态类型区开展迁地保护工作, 同时开展观光木种群生态生殖生物学研究。



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}