



与植物地理学研究相比,菌物地理学研究起步晚,且进展缓慢。早在1792年,德国植物学家Willdenow在其著作Grundlehren der Krauterkunde中,就已经明确提出了植物分布规律的概念,1708年德国学者范·洪堡德(Alexander von Humbolt)出版了Ideen zu einer Géographie der Pflanzen (植物地理学概论),标志着现代地理学的创立(赵万义 2017)。经过200多年的发展,目前植物地理学已经发展成许多分支学科,如植物区系学、区系植物地理学、科属植物地理学、植物分布区学、生态植物地理学及历史植物地理学;建立了完善的学科理论体系和学说,如迁移扩散学说、隔离分化学说;大量相关论著陆续出版,如苏联学者吴鲁夫(E.B. Wulf)出版的《历史植物地理学》(1944)、美国学者古德(R. Good)的The Geography of the Flowering Plants (有花植物地理学) (1953)和苏联学者塔赫他间(A. Takhatajan)的Flowering Plants, Origin and Distpersal (有花植物的起源和散布) (1972);世界植物区系区划方案已经建立,植物区系的基本特征和分区框架也已经完成(吴征镒 1979;吴征镒等 2011;孙航等 2017;赵万义 2017)。动物的生物地理学也建立了相应的学科理论体系,早在1876年已有相关论著出版(雷富民等 2021;李俊洁和黄晓磊 2022)。
反观菌物生物地理学研究,目前还未形成系统的学科理论体系,也未见有相关论著出版。最初人们对真菌的认识,主要通过蘑菇(大型真菌的子实体)。然而,大型真菌不像植物那样一直都长在固定位置,而是在完成生殖过程后,迅速“消失”,子实体什么时候再出现,在哪里出现,这都是难以确定的。此外,在大型真菌研究中,“个体”的概念或者说个体的呈现形式与动植物不同。在动植物中,一般一个受精卵(种子)可以发育成一个独立的、具相对稳定形态结构的个体。然而对于丝状真菌而言,一个孢子萌发后可以形成庞大的菌丝网络,个体在营养阶段以菌丝(体)存在;对于异宗配合的真菌,需要来自不同个体的菌丝通过锁状联合进行核配,才能完成有性生殖,形成子实体。我们见到的蘑菇(子实体)并不是一个独立的个体,而是“果实”。真菌个体的概念,实际上是比较抽象的、没有固定形态的。早期对真菌物种的鉴定主要依据子实体形态特征,然而真菌的形态特征并不稳定,物种的界定十分困难,使得当时的真菌学家们不愿意基于已有的物种分布信息提出物种的分布区,可以说真菌生物地理学研究在20世纪以前的进展是十分缓慢的(Rehead 1989;Lumbsch et al. 2008)。
目前菌物生物地理学研究的热点主要集中于基于分子钟理论的历史生物地理学分析(Hibbett 2001;Matheny et al. 2009;Skrede et al. 2011;Bonito et al. 2013;Cai et al. 2014;He et al. 2017;Zhu et al. 2019),关于菌物区系地理学的研究主要集中于区系的组成及其特点分析(图力古尔和李玉 2000;宋斌和邓旺秋 2001;周绪申等 2010;周丽伟和戴玉成2013;员瑗等 2015;宋斌等 2018;王妍等 2021;武英达等2021),较少关注区系特有现象和区系替代现象。不同菌物类群的演化历史各异,一些适用于动植物的生物地理学理论还未在菌物中得到验证;长距离扩散和隔离分化是解释菌物分布样式和谱系分化的主要理论(韩利红和杨祝良 2020)。
广西邻近东南亚地区,其中,广西的喀斯特地貌是全球最独特的地貌类型之一,孕育着丰富的生物多样性资源。然而,该区域的大型真菌区系调查一直以来没有得到足够的重视,大型真菌多样性和区系情况尚不清楚;同时,石漠化问题严重威胁着该区域的大型真菌多样性,迫切需要对该区域开展大型真菌区系调查和相关的研究。
1 材料与方法
1.1 研究区域自然地理概况
广西喀斯特地貌主要分布于广西的西南部、西北部、中部和东北部,但仍保存有完好喀斯特原始森林的区域主要分布于广西西南部和西北部,该区域也是全国生物多样性保护优先区。本研究选择的广西喀斯特森林代表性区域为:弄岗国家级自然保护区、木论国家级自然保护区和雅长兰科植物国家级自然保护区(以下简称弄岗保护区、木论保护区和雅长保护区)。从地理位置和气候的角度看,这3个保护区位于我国西南部的广西境内,为热带、亚热带季风气候区。
其中,弄岗保护区位于桂西南喀斯特地区,为中越边境地区,在行政区划中归属于崇左市龙州县和宁明县(106°42′28″-107°04′54″ E,22°13′56″- 22°33′09″ N),总面积为10 077 ha;为北热带季节性雨林气候区,气候特点为春夏湿热多雨,夏秋炎热且雨量充沛,冬春气温偏低,干燥少雨;总的来说,“长夏无冬,偶有奇寒”;年平均气温20.8-22.4 ℃,最冷月平均气温通常在13 ℃以上,年最高气温37-39 ℃;年均降雨量1 150- 1 550 mm,最大降水量达2 043 mm,最小为890 mm,降雨集中在5-9月(农绍勤 1988)。
木论保护区位于桂西北喀斯特地区,在行政区划中归属于河池市环江毛南族自治县(107°54′01″-108°05′51″ E,25°07′01″-25°12′22″ N),与贵州茂兰国家级自然保护区相连,总面积10 829.7 ha;为中亚热带季风气候区,该区年平均气温15.0-18.7 ℃,1月平均气温3.4-8.7 ℃,7月平均气温23.0-26.7 ℃;年均降水量为1 530-1 820 mm,雨季(4-8月)降水量为1 207.1 mm,占全年降水量的73.7%,旱季(9月-次年3月)降水量为430.7 mm,占全年的23.3%;林内阴湿,相对湿度大,一般在80%-90% (郑颖吾等 1999)。
雅长保护区位于桂西偏北的喀斯特地区,在行政区划中归属于百色市乐业县(106°11′31″- 106°27′04″ E,24°44′16″-24°53′58″ N),总面积22 062 ha,约为木论保护区或弄岗保护区面积的2倍,为中亚热带季风气候区;深受季风环流和焚风效应的影响,夏季盛行海洋湿润气团,冬季盛行大陆寒冷气团;由于该区山势高大,峰峦连绵、高耸陡峻、地形复杂,形成独特的中亚热带山地气候;年平均气温16.8-21.2 ℃,年平均最高气温23.3-26.9 ℃,年平均最低气温变动12.7-16.0 ℃;气温垂直变化随海拔高度的升高呈递减趋势,但不同高度段的递减率存在差异,海拔600-985 m高度段存在逆温现象,夏季的缓热降温与在冬季的缓冷保温调节作用较明显;年平均降水量940.8-1 216.9 mm,呈现夏季(53.5%- 57.2%)>春季(18.7%-21.4%)>秋季(17.6%-19.7%)> 冬季(4.0%-6.2%)的规律,雨量的季节分配不均匀,春秋干旱,尤其是春旱最为严重(广西林业勘探设计院 2007)。
从植被的组成来看,这3个保护区各具特色。弄岗保护区为北热带季节性雨林区,阔叶林占绝对优势,泛热带分布的植物丰富,如榕属Ficus、风吹楠属Horsfieldia、藤黄属Garcinia、蚬木属Excentrodendron、木奶果属Baccaurea、台湾山柚属Champereia和桄榔属Arenga等,壳斗科Fagaceae和松科Pinaceae植物较少,主要分布于山顶;弄岗保护区的区域特色植物主要有大叶风吹楠Horsfieldia kingii、望天树Parashorea chinensis、董棕Caryota obtusa、桄榔Arenga westerhoutii和蚬木Excentrodendron tonkinense等。
木论保护区植被以中亚热带常绿落叶阔叶混交林为主,山顶为针阔混交林;壳斗科植物种类较多,但数量较少,呈零星分布,多见于山顶;松科植物在木论保护区呈局部分布,山顶较丰富;木论保护区的区域特色植物有掌叶木Handeliodendron bodinieri、伞花木Eurycorymbus cavaleriei、岩生翠柏Calocedrus rupestris和越南黄金柏Xanthocyparis vietnamensis等。与弄岗保护区相比,木论保护区有较丰富的壳斗科和松科等与真菌形成广泛共生关系的植物。
雅长保护区与弄岗保护区和木论保护区的植被相比,最显著的特征为具有大量的、成片分布的兰科Orchidaceae、壳斗科、松科和桦木科Betulaceae等可与真菌形成广泛共生关系的植物类群,使得雅长保护区盛产牛肝菌科Boletaceae、红菇科Russulaceae、鹅膏科Amanitaceae和丝膜菌科Cortinariaceae等外生菌根真菌。
1.2 研究材料
本研究所采用的材料为笔者于2017年5月- 2021年8月在广西弄岗保护区、雅长保护区和木论保护区所采集的大型真菌标本,共计3 800份。其中,雅长保护区收集到1 733份,木论保护区收集到1 147份,弄岗保护区收集到920份。研究的凭证标本保存于吉林农业大学菌物标本馆(HMJAU)以及广西植物标本馆(IBK)。
1.3 研究方法
1.3.1 野外调查和采集
为了尽可能多地获取标本,本研究采用随机取样的方法进行调查取样(Prayudi et al. 2019)。在广西,每个国家级自然保护区都安排有护林员巡护,其巡护线路覆盖整个保护区的主要区域,涵盖不同的生境类型。在大型真菌生长季节(4-11月),按弄岗保护区、木论保护区和雅长保护区的护林员巡护线路及周边范围内开展踏查采集。采集过程中,详细记录每份标本的宏观特征、质地、颜色、气味及伤时颜色变化情况等;伞菌类着重记录菌盖的形态、菌盖条纹有无、菌褶的着生情况、菌柄、菌幕、菌环、菌托和鳞片有无及其特点、菌褶是否自溶等;生态习性如粪生、木生或土生等。多角度拍摄每个物种的生境照片并编号。
1.3.2 标本鉴定
1.3.3 优势科属种的确定
依据区系物种名录,对大型真菌的科、属、种用Excel软件进行归类统计,按各类群物种的数量排序,并综合考虑某物种(类群)采集到的频率,确定优势科、属、种。
1.3.4 地理成分划分
1.3.5 表征科和表征属的确定
综合考虑广西不同气候带喀斯特林区大型真菌非广布成分科和属的数量及在研究区域内的丰富度情况,确定表征科和表征属。
2 结果与分析
2.1 广西喀斯特林区大型真菌区系特点
2.1.1 广西喀斯特林区大型真菌区系的组成
图1

图1
中国新记录种(Ⅰ) A-D:史丹皮尔斯拟针刺菌Mycorrhaphoides stalpersii. 标尺:A,D=5 cm;B=2 cm;C=1 cm. E-F:印度藓菇Rickenella indica. 标尺=5 mm. G-H:假根状线虫草Ophiocordyceps pseudorhizoidea. 标尺=1 cm
Fig. 1
Newly recorded species (Ⅰ). A-D: Mycorrhaphoides stalpersii. Bars: A, D=5 cm; B=2 cm; C=1 cm. E-F: Rickenella indica. Bars=5 mm. G-H: Ophiocordyceps pseudorhizoidea. Bars=1 cm.
图2

图2
中国新记录种(Ⅱ) A-C:东方橘黄裸伞. 标尺=5 cm. D-E:橄榄绿乳菇. 标尺=2 cm. F-G:米色光柄菇. 标尺=2 cm. H-K:异色光柄菇. 标尺=2 cm. L-M:隔囊光柄菇. 标尺=2 cm. N-O:莫洛佐娃草菇. 标尺=5 cm. P-R:棕盖草菇. 标尺=2 cm. S-U:暹罗黄蘑菇. 标尺=2 cm. V-X:玫红靴耳. 标尺V-W=2 cm,X=5 mm
Fig. 2
Newly recorded species (Ⅱ). A-C: Gymnopilus orientispectabilis. Bars=5 cm. D-E: Lactarius olivaceopallidus. Bars=2 cm. F-G: Pluteus flavidus. Bars=2 cm. H-K: Pluteus varius. Bars=2 cm. L-M: Pluteus septocystidiatus. Bars=2 cm. N-O: Volvariella morozovae. Bars=5 cm. P-R: Volvariella pulla. Bars=2 cm. S-U: Xanthagaricus siamensis. Bars=2 cm. V-X: Crepidotus boninensis. Bars: V-W=2 cm, X=5 mm.
表1 广西喀斯特林区大型真菌区系组成
Table 1
| 分类群 Taxon | 纲数 Number of classes | 目数 Number of orders | 科数 Number of families | 属数 Number of genera | 种数 Number of species |
|---|---|---|---|---|---|
| 子囊菌门 Ascomycota | 3 | 5 | 18 | 31 | 74 |
| 担子菌门 Basidiomycota | 3 | 16 | 79 | 287 | 703 |
| 合计 Total amount | 6 | 21 | 97 | 318 | 777 |
未包含科级分类地位未定类群;马勃属Lycoperdon和秃马勃属Calvatia归入蘑菇科Agaricaceae,其余类群参考Index fungorum
The Incertae sedis group not included in this Table; Calvatia and Lycoperdon were included in Agaricaceae, the rest refer to Index fungorum.
2.1.2 科的区系特点
(1) 科的区系组成
在研究区域采集到的大型真菌中,中等科(具有10-30种的科)、较大科(具有31-50种的科)和大科(多于50种的科)共有20个科,占总科数的20.61%,这些科所包含的种数占总种数的比例高达67.39%;寡种科共有51个,占总科数的52.58%,所含的种数占总种数的29.11%;在研究区域只采到1个种的科共有26个,占总科数的26.80%,仅占总种数的3.50%。由此可见,中等科、较大科和大科是构成研究区域大型真菌区系的主要成分(表2)。
表2 广西喀斯特林区大型真菌科的大小分级统计(种级)
Table 2
| 级别 Grade | 科数 Number of families | 占总科数的百分比 Proportion to total Families (%) | 种数 Number of species | 占总种数的百分比 Proportion to total species (%) |
|---|---|---|---|---|
| 大科(>50种) Fam. of more than 50 spec. collected | 2 | 2.06 | 146 | 19.68 |
| 较大科(31-50种) Fam. of 31-50 spec. collected | 2 | 2.06 | 90 | 12.13 |
| 中等科(10-30种) Fam. of 10-30 spec. collected | 16 | 16.49 | 264 | 35.58 |
| 寡种科(2-9种) Fam. of 2-9 spec. collected | 51 | 52.58 | 216 | 29.11 |
| 仅采到1种的科 Fam. of only 1 spec. collected | 26 | 26.80 | 26 | 3.50 |
科级分类地位未定的类群未统计在内;本表统计的总科数为97个,总种数为742种
The incertae sedis group not included in this Table; Only 97 families and 742 species were included in this Table.
其中,大科(多于50种的科)有2个:多孔菌科Polyporaceae (86种)和蘑菇科Agaricaceae (60种)。这两个科均为世界广布科,虽然仅占总科数的2.06%,却占总种数的19.68%,在广西喀斯特林区大型真菌区系中具有重要地位,且为优势类群。
较大科(具31-50种的科)有2个:牛肝菌科Boletaceae (42种)和红菇科Russulaceae (48种)。这2个科所含物种数占总种数的12.13%。牛肝菌科和红菇科均为世界广布科,且为外生菌根真菌,在研究区域内主要分布于雅长保护区;木论保护区有少量分布,在弄岗保护区未发现有这2个科的物种。这2个科的物种与壳斗科、松科和桦木科等植物类群形成共生关系,一个地区这些植物的多寡直接影响到这2个科真菌的数量。从弄岗保护区、木论保护区和雅长保护区内这些植物类群的多寡来看,牛肝菌科和红菇科物种在这些保护区的数量和分布情况是基本一致的。然而,在弄岗保护区内有壳斗科和松科的植物,却未发现有牛肝菌科及红菇科的物种,这可能和壳斗科、松科植物在弄岗保护区内的分布有关。壳斗科、松科植物在弄岗保护区内主要分布于山顶,由于弄岗保护区地处于热带雨林区,淋溶作用强烈,导致山顶无法积淀腐殖质层,为裸露的岩石。牛肝菌科和红菇科真菌为土生真菌,缺乏土壤直接影响这些科物种的生长发育,也就难以产生子实体,故在调查中未在弄岗保护区发现牛肝菌科和红菇科等外生菌根真菌。
中等科(具10-30种的科)有16个:肉杯菌科Sarcoscyphaceae (14种)、炭团菌科Hypoxylaceae (12种)、炭角菌科Xylariaceae (20种)、鹅膏科Amanitaceae (17种)、丝膜菌科Cortinariaceae (21种)、靴耳科Crepidotaceae (17种)、层腹菌科Hymenogastraceae (16种)、丝盖伞科Inocybaceae (12种)、小皮伞科Marasmiaceae (21种)、小菇科Mycenaceae (16种)、类脐菇科Omphalotaceae (21种)、膨瑚菌科Physalacriaceae (12种)、光柄菇科Pluteaceae (10种)、小脆柄菇科Psathyrellaceae (16种)、木耳科Auriculariaceae (13种)和锈革菌科Hymenochaetaceae (26种)。中等科占总科数的16.49%,科内种数占总种数的35.58%,是广西喀斯特林区大型真菌区系的重要数量优势科。其中,肉杯菌科、炭角菌科、小皮伞科、小菇科和光柄菇科为泛热带成分科,所含物种数(81种)占非广布成分总种数(213种)的38.03%,是反映热带区系性质的重要科。中等科中,属于温带成分的科仅有丝膜菌科(1属,21种),这也从侧面反映出广西喀斯特林区的大型真菌区系性质为热带亚热带区系性质。丝膜菌科的物种也为外生菌根真菌,也仅见于木论和雅长保护区。
寡种科(具2-9种的科)有51个:裂皮盘菌科Chorioactidaceae、火丝菌科Pyronemataceae、麦角菌科Clavicipitaceae、虫草菌科Cordycipitaceae、肉座菌科Hypocreaceae、线虫草科Ophiocordycipitaceae、角孢伞科Asproinocybaceae、粪锈伞科Bolbitiaceae、珊瑚菌科Clavariaceae、粉褶菌科Entolomataceae、轴腹菌科Hydnangiaceae、蜡伞科Hygrophoraceae、离褶伞科Lyophyllaceae、侧耳科Pleurotaceae、木须菌科Lentariaceae、羽瑚菌科Pterulaceae、裂褶菌科Schizophyllaceae、球盖菇科Strophariaceae、口蘑科Tricholomataceae、假脐菇科Tubariaceae、圆孔牛肝菌科Gyroporaceae、铆钉菇科Gomphidiaceae、桩菇科Paxillaceae、硬皮马勃科Sclerodermataceae、乳牛肝菌科Suillaceae、齿菌科Hydnaceae、地星科Geastraceae、鬼笔科Phallaceae、褐褶菌科Gloeophyllaceae、藓菇科Rickenellaceae、裂孔菌科Schizoporaceae、钉菇科Gomphaceae、下皮黑孔菌科Cerrenaceae、拟层孔菌科Fomitopsidaceae、硫磺菌科Laetiporaceae、灵芝科Ganodermaceae、蜡孢菌科Incrustoporiaceae、囊耙齿菌科Irpicaceae、干朽菌科Meruliaceae、锐孔菌科Oxyporaceae、革耳科(暂定名) Panaceae、原毛平革菌科Phanerochaetaceae、柄杯菌科Podoscyphaceae、齿耳科Steccherinaceae、耳匙菌科Auriscalpiaceae、刺孢多孔菌科Bondarzewiaceae、韧革菌科Stereaceae、坂氏齿菌科Bankeraceae、革菌科 Thelephoraceae、银耳科Tremellaceae和花耳科Dacrymycetaceae。寡种科在总科数中的占比超过一半,但所含的物种数仅占总种数的29.11%。其中,广布成分科有32个,热带成分科有8个(39种),温带成分科有11个(57种)。寡种科中,热带成分总科数和种数分别占研究区域热带成分总科数的40%,总种数的30.71%;温带成分科数和种数分别占研究区域温带成分总科数的55%,总种数的66.28%。这说明寡种科是反映热带区系性质的重要来源科,反映科温带性质的主要来源科。
在研究区域只采到1个种的科有26个:绿杯盘菌科Chlorociboriaceae、复柄盘菌科(暂定名) Cordieritidaceae、柔膜菌科Helotiaceae、粒毛盘菌科Lachnaceae、蜡盘菌科Rutstroemiaceae、锤舌菌科Leotiaceae、垫盘菌科(暂定名) Pulvinulaceae、马鞍菌科Helvellaceae、肉盘菌科Sarcosomataceae、Biannulariaceae、色孢菌科Callistosporiaceae、囊韧革菌科Cystostereaceae、核瑚菌科Typhulaceae、淀粉伏革菌科Amylocorticiaceae、小牛肝菌科Boletinellaceae、丽口菌科Calostomataceae、粉孢革菌科Coniophoraceae、复囊菌科Diplocystidiaceae、干腐菌科Serpulaceae、点革菌科(暂定名) Punctulariaceae、索孔菌科Fibroporiaceae、地花孔菌科Albatrellaceae、猴头菌科Hericiaceae、蜡壳耳科Sebacinaceae、拟韧革菌科Stereopsidaceae和胶珊瑚科Holtermanniaceae。这些科中,广布成分科有12个,热带成分科有7个,温带成分科有8个;热带成分总科数和种数分别占研究区域热带成分总科数的35%,总种数的5.51%;温带成分科数和种数分别占研究区域温带成分总科数的40%,总种数的9.30%,说明研究区域内只发现1种的科是反映科区系性质的重要来源科,但不是优势科。
从科内属级的分析(表3)来看,研究区域分布多于30个属的科为多孔菌科,有40属86种,占总科数的1.03%、总属数的13.38%、总种数的11.59%。含21-30个属的科有牛肝菌科(21属,42种),占总科数的1.03%、总属数的7.02%、总种数的5.66%。具10-20个属的科有蘑菇科(10属,60种)和锈革菌科(13属,26种),合计占总科数的2.06%、总属数的7.69%、总种数的11.59%。
表3 广西喀斯特林区大型真菌科的大小分级统计(属级)
Table 3
| 级别 Grade | 科数 Number of families | 占总科数的 百分比 Proportion to total families (%) | 属数 Number of genera | 占总属数的 百分比 Proportion to total genera (%) | 种数 Number of species | 占总种数的 百分比 Proportion to total species (%) |
|---|---|---|---|---|---|---|
| >30个属的科 Fam. of more than 30 gen. collected | 1 | 1.03 | 40 | 13.38 | 86 | 11.59 |
| 21-30个属的科 Fam. of 21-30 gen. collected | 1 | 1.03 | 21 | 7.02 | 42 | 5.66 |
| 10-20个属的科 Fam. of 10-20 gen. collected | 2 | 2.06 | 23 | 7.69 | 86 | 11.59 |
| 2-9个属的科 Fam. of 2-9 gen. collected | 52 | 53.61 | 174 | 58.19 | 448 | 60.38 |
| 仅采到1个属的科 Fam. of only 1 gen. collected | 41 | 42.27 | 41 | 13.71 | 80 | 10.78 |
科级分类地位未定的类群没有统计在内;本表统计的总科数为97个,总属数为299个,总种数为742个
The incertae sedis group not included in this Table; Only 97 families, 299 genera and 742 species were included in this Table.
科内所含属数有10个属及以上的科均为世界广布科,以多孔菌科所含的属数和种数都最多。广西喀斯特林区的典型特征为岩石裸露度高(可达90%),土层及腐殖质层稀薄,不利于土生真菌的生长;虽然岩石裸露度高,植被却十分茂密,产生大量的枯枝腐木,为木生真菌生长提供了有利条件,因而木生真菌为广西喀斯特森林大型真菌的优势类群,属数大于10个的科主要来自木腐菌。牛肝菌科真菌所含的属数和种数也较为丰富,主要分布于壳斗科和松科植物丰富的雅长保护区。
科内具2-9个属的科有52个,占总科数的53.61%,占总属数的58.19%,占总种数的60.38%。具有2-9个属的科中,广布成分的科有29个,热带成分的科有12个,温带成分的科有10个;热带成分的科数及科内属数、种数分别占研究区域内热带成分总科数的60%,总属数的82.98%,总种数的94.49%;温带成分的科数及科内属数、种数分别占研究区域内温带成分总科数的50%,总属数的75%,总种数的66.28%。说明含2-9个属的科为反映广西喀斯特林区大型真菌区系的主要来源科。
仅采集到1个属的科共有41个,占总科数的42.27%、总属数的13.71%、总种数的10.78%。仅采集到1个属的科中,广布成分的科有24个,热带成分科有8个,温带成分科有10个;这些科除了丝膜菌科的丝膜菌属含21种,其余科均为寡种科、寡种属,说明仅含1个属的科不是优势科,但是反映了广西喀斯特林区大型真菌区系的重要来源科。
(2) 科的区系成分
由于当前大型真菌科级水平的分布区划尚未厘清,也未形成统一的区划认识,本研究仅简单将其划分为广布成分、泛热带成分和北温带成分。
根据划分结果(表4),广西喀斯特林区大型真菌科的分布类型以广布成分为主(58.16%),泛热带成分(20.41%)和北温带成分(21.43%)具有相近的比例。
表4 广西喀斯特林区大型真菌科的分布区类型统计
Table 4
| 分布区类型 Distribution-type | 科数 Number of families | 占总科数的百分比 Proportion to total families (%) | 占非世界分布科的百分比 Proportion to non-cosmopolitan families (%) |
|---|---|---|---|
| 广布成分 Cosmopolitan | 57 | 58.16 | - |
| 泛热带成分 Pantropic | 20 | 20.41 | 48.78 |
| 北温带成分 N. Temp. | 21 | 21.43 | 51.22 |
泛热带成分的科有:小牛肝菌科(1属,1种)、丽口菌科(1属,1种)、麦角菌科(1属,2种)、粉褶菌科(2属,5种)、灵芝科(2属,3种)、柔膜菌科(1属,1种)、胶珊瑚科(1属,1种)、小皮伞科(5属,21种)、小菇科(6属,16种)、革耳科(2属,4种)、鬼笔科(6属,8种)、光柄菇科(2属,10种)、柄杯菌科(2属,6种)、羽瑚菌科(3属,5种)、点革菌科(1属,1种)、肉杯菌科(4属,14种)、肉盘菌科(1属,1种)、拟韧革菌科(1属,1种)、银耳科(2属,6种)和炭角菌科(3属,20种)。泛热带成分的科中,在弄岗保护区有分布的16个,其中小牛肝菌科、麦角菌科和拟韧革菌科仅见于弄岗保护区;在雅长保护区有分布的16个,其中丽口菌科和胶珊瑚科仅见于雅长保护区;在木论保护区有分布的科 12个,肉盘菌科仅见于木论保护区。
北温带成分的科有:锤舌菌科(1属,1种)、地花菌科(1属,1种)、淀粉伏革菌科(1属,1种)、坂氏齿菌科(2属,3种)、刺孢多孔菌科(3属,3种)、复柄盘菌科(1属,1种)、丝膜菌科(1属,21种)、索孔菌科(1属,1种)、拟层孔菌科(6属,7种)、褐褶菌科(2属,2种)、钉菇科(4属,9种)、铆钉菇科(2属,2种)、马鞍菌科(1属,1种)、猴头菌科(1属,1种)、齿菌科(3属,8种)、蜡伞科(3属,6种)、离褶伞科(3属,9种)、干腐菌科(1属,1种)、乳牛肝菌科(1属,3种)、革菌科(2属,5种)和核瑚菌科(1属,1种)。北温带成分的科中,分布于弄岗保护区的仅3个;分布于雅长保护区的有18个,其中锤舌菌科、地花菌科、淀粉伏革菌科、索孔菌科、褐褶菌科、齿菌科、蜡伞科和革菌科仅见于雅长保护区;分布于木论保护区的有13个,其中干腐菌科和核瑚菌科仅见于木论保护区。
考虑到本研究实际上选择的是不同纬度带的区域进行研究,可能按不同地区分别来统计更能体现实际的区系情况。分类统计过程中,用热带成分科数与北温带成分科数的比值来反映一个地区科的区系性质。当这个比值大于1时,说明这个地区科的区系成分为偏热带性质;当比值小于1时,说明为偏温带性质。
统计结果(表5)表明,雅长保护区科的区系成分偏温带性质,弄岗保护区为明显的热带性质,木论保护区则呈现热带和温带过渡,这个结果比较符合实际情况。雅长保护区和木论保护区都位于桂西北地区且在纬度上相差不大,但是雅长保护区所在地海拔相对较高,海拔1 000 m以上的山峰共有89座,1 500 m以上的有19座,平均海拔在1 000 m以上;木论保护区的海拔多为400-600 m,海拔上的差异使得雅长保护区比木论保护区表现出更明显的温带区系性质。弄岗保护区位于热带北缘,科的热带性质明显,符合该地区的气候特点。
表5 不同保护区大型真菌科的分布区类型统计
Table 5
| 采集地 Location | 广布成分科数 Cosmopolitan families | 泛热带成分科数 Pantropic families | 北温带成分科数 N. Temp. families | 泛热带成分/北温带成分 Pantropic/N. Temp. |
|---|---|---|---|---|
| 雅长保护区 Yachang Nat. Res. | 52 | 15 | 18 | 0.83 |
| 木论保护区 Mulun Nat. Res. | 44 | 13 | 13 | 1 |
| 弄岗保护区 Nonggang Nat. Res. | 34 | 16 | 3 | 5.3 |
从非广布成分科在弄岗、木论、雅长保护区的分布情况来看,雅长保护区不仅具有较多的温带成分科,还具有和弄岗保护区相同的热带成分科数,这说明雅长保护区虽然整体上表现出偏温带区系性质,但热带成分科也被较好地保存,可见其区系之复杂,这与雅长保护区位于我国阶梯地势第二级与第三级的过渡地区,也是热带向亚热带过渡的地区,以及亚热带常绿阔叶林带东部湿润区与西部半湿润区的交接地带的特殊地理位置有关。
(3) 优势科
优势科是指在区系成分中所包含的属、种相对较多的科。通过前面科内属、种的数量结构分析,知道多孔菌科(40属,86种)、蘑菇科(10属,60种)、牛肝菌科(21属,42种)、红菇科(3属,48种)、锈革菌科(13属,26种)、类脐菇科(7属,21种)、丝膜菌科(1属,21种)、小皮伞科(5属,21种)、炭角菌科(3属,20种)、小菇科(6属,16种)、炭团菌科(5属,12种)、膨瑚菌科(9属,12种)、层腹菌科(5属,16种)和小脆柄菇科(4属,16种)所包含的属数或种数较多。
综合考虑这些科的属、种数量以及科内物种在研究区域内出现的频率,本研究将多孔菌科、牛肝菌科、蘑菇科、锈革菌科、类脐菇科、小皮伞科、小菇科、膨瑚菌科、炭团菌科、层腹菌科和小脆柄菇科确定为广西喀斯特林区的优势科。值得说明的是,虽然牛肝菌科所含的属数(21属)和种数(42种)都很多,但该科主要见于雅长保护区,目前在弄岗保护区还未发现有该科物种的分布,该科物种在木论保护区内也不十分常见。这11个优势科占总科数的11.34%,所含的属数和种数分别占总属数和总种数的39.30%和42.21%。这些优势科大部分为木生真菌,符合广西喀斯特森林的生态特点。
(4) 表征科
表征科是指那些能代表真菌区系的科,所含的属、种相对非广布成分占有较大比重。通过前面科的地理成分分析,知道非广布成分的科中,小皮伞科(5属,21种)、小菇科(6属,16种)、鬼笔科(6属,8种)、光柄菇科(2属,10种)、柄杯菌科(2属,6种)、羽瑚菌科(3属,5种)、肉杯菌科(4属,14种)、银耳科(2属,6种)和炭角菌科(3属,20种)这些泛热带分布的科;以及丝膜菌科(1属,21种)、拟层孔菌科(6属,7种)、钉菇科(4属,9种)、齿菌科(3属,8种)、蜡伞科(3属,6种)、离褶伞科(3属,9种)和革菌科(2属,5种)这些北温带的科所含的属数、种数较多。
综合考虑这些科的属、种数量以及科内物种在研究区域内出现的频率,本研究将小皮伞科、小菇科、鬼笔科、柄杯菌科、肉杯菌科、银耳科和炭角菌科这些泛热带分布的科,以及拟层孔菌科、钉菇科、齿菌科和离褶伞科这些北温带分布的科确定为广西喀斯特地区大型真菌区系的表征科。这11个表征科占非广布成分科总数的27.50%,总属数的50.57%,总种数的58.22%。表征科中,泛热带成分科数占非广布成分科总数的17.50%,总属数的32.18%,总种数的42.72%;北温带成分科数占非广布成分科总数的10.00%,总属数的18.39%,总种数的15.50%。广西喀斯特林区表征科同样为木生型真菌明显占优势。
2.1.3 属的区系特点
(1) 属的区系组成
通过分析广西不同纬度带喀斯特林区的3个具代表性的国家级自然保护区的大型真菌区系属、种的数量,对属的大小进行分级,结果见表6。
表6 广西喀斯特林区大型真菌属的大小分级统计
Table 6
| 级别 Grade | 子囊菌的属数 Number of genera in Ascomycota | 担子菌的属数 Number of genera in Basidiomycota | 属合计 Number of total genera | 占总属数百分比 Proportion to total genera (%) | 种数 Number of species | 占总种数的百分比 Proportion to total species (%) |
|---|---|---|---|---|---|---|
| 大属(>20种) Gen. of >20 spec. | - | 3 | 3 | 0.94 | 64 | 8.24 |
| 较大属(10-20种) Gen. of 10-20 spec. | 1 | 8 | 9 | 2.83 | 116 | 14.93 |
| 中等属(5-9种) Gen. of 5-9 spec. | 2 | 23 | 25 | 7.86 | 144 | 18.53 |
| 寡种属(2-4种) Gen. of 2-4 spec. | 11 | 97 | 108 | 34.28 | 280 | 36.03 |
| 仅采到1个种的属 Gen. of only 1 spec. | 17 | 156 | 173 | 54.40 | 173 | 22.27 |
研究区域所有真菌类群中,具有种数超过20种的属(大属)有3个:丝膜菌属Cortinarius (21种)、乳菇属Lactarius (21种)和红菇属Russula (22种),占总属数的0.94%、总种数的8.24%。
具10-20种的属(较大属)有9个:炭角菌属Xylaria (18种)、蘑菇属Agaricus (15种)、白环蘑属Leucoagaricus (12种)、鹅膏属Amanita (16种)、靴耳属Crepidotus (11种)、丝盖伞属Inocybe (10种)、小皮伞属Marasmius (14种)、裸脚伞属Gymnopus (10种)和栓孔菌属Trametes (10种),占总属数的2.83%、总种数的14.93%。
具5-9种的属(中等属)有25个:毛杯菌属Cookeina (5种)、线虫草属Ophiocordyceps (5种)、老伞属Gerronema (5种)、环柄菇属Lepiota (9种)、马勃属Lycoperdon (6种)、小蘑菇属Micropsalliota (6种)、绒盖伞属Simocybe (5种)、黄囊菇属Deconica (5种)、蚁巢伞属Termitomyces (7种)、小菇属Mycena (7种)、微皮伞属Marasmiellus (5种)、光柄菇属Pluteus (6种)、小鬼伞属Coprinellus (5种)、拟鬼伞属Coprinopsis (5种)、小脆柄菇属Psathyrella (5种)、木耳属Auricularia (8种)、锈革菌属Hymenochaete (7种)、枝瑚菌属Ramaria (6种)、柄杯菌属Podoscypha (5种)、棱孔菌属Favolus (5种)、灵芝属Ganoderma (7种)、多孔菌属Polyporus (5种)、多汁乳菇属Lactifluus (5种)、革菌属Thelephora (5种)和银耳属Tremella (5种),占总属数的7.86%、总种数的18.53%。
具2-4种以及仅具1种的属分别有108个属(284种)和173个属(种),分别占总属数的34.28%和54.40%,分别占总种数的36.03%和22.27%,为研究区域大型真菌的主要组成属。
(2) 属的区系成分
表7 广西喀斯特林区大型真菌属的分布区类型统计
Table 7
| 分布带 Distribution zone | 分布区类型 Distribution-type | 属数 Number of genera | 占非广布成分比例 Proportion to non-cosmopolitan (%) |
|---|---|---|---|
| 广布属(173个) Wide spread | 1. 广布成分Cosmopolitan | 173 | - |
| 热带属(88个) Tropical | 2. 泛热带成分Pantropic | 75 | 51.72 |
| 3. 热带亚洲-热带美洲成分 Trop. As. and Trop. Amer. Disjuncted | 4 | 2.76 | |
| 4. 热带亚洲-热带非洲成分 Trop. As. and Trop. Afr. Disjuncted | 2 | 1.38 | |
| 5. 古(旧)热带成分Old World Trop. | 3 | 2.07 | |
| 6. 热带亚洲成分Trop. Asia | 1 | 0.69 | |
| 7. 热带亚洲至热带大洋洲成分 Trop. Asia, Austalasia Disjuncted | 1 | 0.69 | |
| 8. 热带亚洲、大洋洲至热带美洲成分 Trop. Asia, Austalasia & Amer. Disjuncted | 3 | 2.07 | |
| 温带属(51个) Temperate | 9. 北温带成分N. Temp. | 45 | 31.03 |
| 10. 东亚-北美洲成分 E. Asia and N. Amer. Disjuncted | 4 | 2.76 | |
| 11. 东亚成分E. Asia | 2 | 1.38 | |
| 中国特有属(5个) Endemic to China | 12. 中国特有成分Endemic to China | 5 | 3.45 |
(Ⅰ) 世界广布的属
世界广布属为扩散能力较强的属或包含有多种的大属,在世界各地均有分布。广西喀斯特林区的世界广布属共有173个,占研究总属数的54.40%,含456种,占研究总种数的58.69%。在广布属中,所具种数有5种及以上的属有23个:蘑菇属、鹅膏属、木耳属、丝膜菌属、小鬼伞属、拟鬼伞属、靴耳属、黄囊菇属、裸脚伞属、光柄菇属、多孔菌属、小脆柄菇属、枝瑚菌属、红菇属、绒盖伞属、栓孔菌属、银耳属、环柄菇属、白环蘑属、马勃属、微皮伞属、小菇属和线虫草属。这些属占总属数的7.23%,所具有的物种数(205种)占总种数的26.38%,为广西喀斯特林区大型真菌优势类群的来源属。
具2-4种的广布属有62个:小孢盘菌属Acervus、金牛肝菌属Aureoboletus、牛肝菌属Boletus、秃马勃属Calvatia、锁瑚菌属Clavulina、鸡油菌属Cantharellus、下皮黑孔菌属Cerrena、珊瑚菌属Clavaria、拟锁瑚菌属Clavulinopsis、杯伞属Clitocybe、集毛菌属Coltricia、拱顶菇属Cuphophyllus、黑蛋巢菌属Cyathus、环伞属Cyclocybe、轮层炭壳菌属Daldinia、环鳞伞属Descolea、黑耳属Exidia、小嗜蓝孢孔菌属Fomitiporella、嗜蓝孢孔菌属Fomitiporia、拟层孔菌属Fomitopsis、褐卧孔菌属Fuscoporia、盔孢伞属Galerina、地星属Geastrum、圆孔牛肝菌属Gyroporus、裸伞属Gymnopilus、亚侧耳属Hohenbuehelia、炭团菌属Hypoxylon、蜡蘑属Laccaria、㶷孔菌属Laetiporus、木瑚菌属Lentaria、黑斑根孔菌属Picipes、侧耳属Pleurotus、粉末牛肝菌属Pulveroboletus、密孔菌属Pycnoporus、伏褶菌属Resupinatus、藓菇属Rickenella、红牛肝菌属Rubroboletus、肉齿菌属Sarcodon、硬皮马勃属Scleroderma、干皮孔菌属Skeletocutis、韧革菌属Stereum、松塔牛肝菌属Strobilomyces、革菌属Thelephora、银耳属Tremella、木霉属Trichoderma、Vitreoporus、草菇属Volvariella、黄蘑菇属Xanthagaricus、干脐菇属Xeromphalina、趋木菌属Xylobolus、大环柄菇属Macrolepiota、绿僵菌属Metarhizium、黑孔菌属Nigroporus、类脐菇属Omphalotus、锐孔菌属Oxyporus、花褶伞属Panaeolus、扇菇属Panellus、革耳属Panus、多年卧孔菌属Perenniporia、鬼笔属Phallus、射脉革菌属Phlebia和拟射脉菌属Phlebiopsis。这些属占总属数的19.50%,所具物种数(162种)占总种数的20.85%。
仅采到1种的广布属有89个:残孔菌属Abortiporus、环纹炭团菌属Annulohypoxylon、薄孔菌属Antrodia、分钉耳属Aporpium、蜜环菌属Armillaria、耳匙菌属Auriscalpium、硬皮地星属Astraeus、棕孔菌属Brunneoporus、黄肉牛肝菌属Butyriboletus、絮干朽菌属Byssomerulius、丽口菌属Calostoma、平栓菌属Leiotrametes (Cubamyces)、角孔菌属Cerioporus、色孢菌属Callistosporium、胶角耳属Calocera、绿杯盘菌属Chlorociboria、丽齿耳属Climacodon、复柄盘菌属Cordierites、虫草菌属Cordyceps、毛皮伞属Crinipellis、囊韧革菌属Cystidiodontia、花耳属Dacrymyces、迷孔菌属Daedalea、拟迷孔菌属Daedaleopsis、齿伏革菌属Dentocorticium、无环蜜环菌属Desarmillaria、盘革耳属Eichleriella、小绵杯盘菌属Erioscyphella、黄囊孔菌属Flavodon、索孔菌属Fibroporia、黄褐层孔菌属Fulvifomes、褐褶菌属Gloeophyllum、短孢牛肝菌属Gyrodon、黏菇属Gloiocephala、圆齿菌属Gyrodontium、被毛孢属Hirsutella、产丝齿菌属Hyphodontia、Hodophilus、齿菌属Hydnum、漏斗伞属Infundibulicybe、纤孔菌属Inonotus、炭墩菌属Kretzschmaria、垂暮菇属Hypholoma、刺射脉菌属Hydnophlebia、容氏孔菌属Junghuhnia、毡毛脆柄菇属Lacrymaria、兰斯盘菌属Lanzia、松肉菌属Laxitextum、亚卧孔菌属Physisporinus、豆马勃属Pisolithus、翁孔菌属Onnia、暗银耳属Phaeotremella、原毛平革菌Phanerochaete、拟褶尾菌属Plicaturopsis、孔座壳属Poronia、原球盖菇属Protostropharia、裸盖菇属Psilocybe、孔韧革菌属Porostereum、垫盘菌属Pulvinula、点革菌属Punctularia、红层孔菌属Rhodofomes、裂褶菌属Schizophyllum、干腐菌属Serpula、盾盘菌属Scutellinia、蜡壳耳属Sebacina、齿耳菌属Steccherinum、球盖菇属Stropharia、拟栓菌属Trametopsis、红金钱菌属Rhodocollybia、十字孢伞属Tricholosporum、干酪菌属Tyromyces、万德孔菌属Vanderbylia、金孢牛肝菌属Xanthoconium、绒盖牛肝菌属Xerocomus、革裥菌属Lenzites、小香菇属Lentinellus、黏伞属Limacella、齿脉菌属Lopharia,Meiorganum、齿孔菌属Metuloidea、拟干朽菌属Meruliopsis、蛇头菌属Mutinus、蒜味皮伞属Mycetinis、针菌属Mycoacia、新香菇属Neolentinus、红蛋巢菌属Nidula、白孔层孔菌属Niveoporofomes、桩菇属Paxillus和木层孔菌属Phellinus。这些属占总属数的28.00%,所具物种数占总种数的11.45%。
在仅采集到1种的广布属中,既有十分常见的,也有仅发现于某个保护区的。比较典型的有:胶角耳属(角状胶角耳Calocera cornea)、小绵杯盘菌属(异常小绵杯盘菌Erioscyphella abnormis)、裂褶菌属(裂褶菌Schizophyllum commune)、盾盘菌属(盾盘菌Scutellinia scutellata)和木层孔菌属(淡黄木层孔菌Phellinus gilvus)。这些种在调查的不同区域内都十分常见,这些仅发现有1种的属对应的种可能是广布种或泛热带、亚热带分布的种。对于仅发现于某个保护区的属(种),将在区系替代性一文中讨论。值得说明的是,这些目前仅发现有1种的广布属,除了裂褶菌属,其他属并不能说明就是单种属,可能还有其他的同属内的物种在研究区域内没有被发现。
(Ⅱ) 泛热带成分的属
泛热带分布的属是指分布于东、西两半球热带,或可达亚热带至温带,但分布中心仍在热带的属。发现于广西喀斯特林区内泛热带分布的属有75个,占扣除广布成分后总属数的51.72%,体现出广西喀斯特林区大型真菌区系整体情况呈热带性质的特点,符合广西地处热带、亚热带季风气候区的特点。
泛热带成分的属中,属内所具物种数5种及以上的属有9个:毛杯菌属、棱孔菌属、灵芝属、老伞属、炭角菌属、锈革菌属、小皮伞属、小蘑菇属和柄杯菌属。这些属占扣除广布成分后总属数(145属)的6.21%,所具物种数(72种)占扣除广布成分后总种数(320种)的22.5%。这些属不仅是广西喀斯特林区的优势属,也是反映热带区系成分的核心属,在热带北缘的弄岗保护区更为常见。
泛热带成分的属中,属内所具物种数为2-4种的属有31个:条孢牛肝菌属Boletellus、美牛肝菌Caloboletus、毛筐菌属Chaetocalathus、杯革菌属Cotylidia、鳞盖伞属Cyptotrama、青褶伞属Chlorophyllum、棘刚毛菌属Echinochaete、粉褶菌属Entoloma、胶球炭壳菌属Entonaema、胶孔菌属Favolaschia、粗毛盖菌属Funalia、蜂窝孔菌属Hexagonia、香菇属Lentinus、白鬼伞属Leucocoprinus、巨孔菌属Megasporia、小孔菌属Microporus、新大孔菌属Neofavolus、小奥德蘑属Oudemansiella、歪盘菌属Phillipsia、褶孔牛肝菌属Phylloporus、须瑚菌属Pterula、辛格杯伞属Singerocybe、四角孢属Tetrapyrgos、色孔菌属Tinctoporellus、胶陀盘菌属Trichaleurina、附毛孔菌属Trichaptum、热带孔菌属Tropicoporus、畸孢孔菌属Truncospora和趋木齿菌属Xylodon。这些属占扣除广布成分后总属数(145属)的21.38%,所具物种数(73种)占扣除广布成分后总种数(320种)的22.81%。这些属中,除了美牛肝菌、青褶伞属、胶球炭壳菌属和褶孔牛肝菌属这几个属仅见于某个保护区,其他属在广西喀斯特林区内都较为常见,也是反映热带区系成分的重要组成部分。
泛热带成分的属中,属内所发现物种数仅1种的属有37个:粉孢菌属Amylosporus、角孢伞属Asproinocybe、星头鬼笔属Aseroe、炭褶菌属Anthracophyllum、布氏多孔菌属Bresadolia、脉褶菌属Campanella、革孔菌属Coriolopsis、Crassisporus、条孢灵芝属(暂定名) Cristataspora、赤小菇属Cruentomycena、波边革菌属Cymatoderma、假花耳属Dacryopinax、小异薄孔菌属Datroniella、龙爪菌属Deflexula、膜齿菌属Dentipellicula、俄氏孔菌属Earliella、榆孔菌属Elmerina、网孢牛肝菌属Heimioporus、Hirticrusta、锈齿革菌属Hydnochaete、长根菇属Hymenopellis、考巴菌属Kobayasia、乳金伞属Lactocollybia、木菇属Lentinula、散尾鬼笔属Lysurus、小小孔菌属Microporellus、Mycorrhaphoides、暗锁瑚菌属Phaeoclavulina、脉柄牛肝菌属Phlebopus、叶孔菌属Phylloporia、泡头菌属Physalacria、晶星革菌属Sidera、链担耳属Sirobasidium、拟韧革菌属Stereopsis、沟褶菌属Trogia,Trullella和小剥管孔菌属Piptoporellus。这些属占扣除广布成分后总属数(145属)的25.52%,所具物种数(70种)占扣除广布成分后总种数(320种)的11.56%。
(Ⅲ) 热带亚洲-热带美洲成分的属
指间断分布于亚洲和美洲热带地区的属,其中某些属可达这两洲的亚热带地区。该成分的属有4个,属内种数在研究区域内均只发现1种,占扣除广布成分后总属数(145属)的2.76%,占扣除广布成分后总种数(320种)的1.25%。这4个属为:大孢孔菌属Megasporoporia、侧火菇属Pleuroflammula、湿星伞属Hygroaster和皱盖牛肝菌属Rugiboletus。
(Ⅳ) 热带亚洲-热带非洲成分的属
指仅分布于热带亚洲至热带非洲的属。该成分的属有2个,属内种数共9种,占扣除广布成分后总属数(145属)的1.38%,占扣除广布成分后总种数(320种)的2.81%。这2个属为:槌座炭角菌属Rhopalostroma (2种)和蚁巢伞属(7种)。
(Ⅴ) 古(旧)热带成分的属
指间断分布于亚洲、非洲和大洋洲热带地区及其邻近岛屿的属。该成分的属有3个,属内种数共4种,占扣除广布成分后总属数(145属)的2.07%,占扣除广布成分后总种数(320种)的1.25%。这3个属为:血芝属(暂定名) Sanguinoderma (2种)、核生柄孔菌属Lignosus (1种)和淀粉孔菌属Amylonotus (1种)。
(Ⅵ) 热带亚洲成分的属
指主要分布于南亚、东南亚等地,东面可到斐济等太平洋岛屿,但不达澳大利亚大陆,其分布的北缘可达我国西南、华南至台湾,甚至更北地区的属。该成分的属仅有羽囊菌属Pterulicium这1属,具1种,占扣除广布成分后总属数(145属)的0.69%,占扣除广布成分后总种数(320种)的0.31%。
(Ⅶ) 热带亚洲至热带大洋洲成分的属
指分布于热带亚洲至大洋洲旧世界热带的东翼,一般不到非洲大陆的属。该成分的属仅有新小层孔菌属Neofomitella这1属,具2种,占扣除广布成分后总属数(145属)的0.69%,占扣除广布成分后总种数(320种)的0.63%。
(Ⅷ) 热带亚洲、大洋洲至热带美洲成分的属
指间断分布于热带亚洲、大洋洲和美洲的属。该成分的属有3个,共3种,占扣除广布成分后总属数(145属)的2.07%,占扣除广布成分后总种数(320种)的0.94%。这3个属为:二头孢盘菌属Dicephalospora (1种)、盘孔菌属Porodisculus (1种)和胶珊瑚属Holtermannia (1种)。
(Ⅸ) 北温带成分的属
指分布于北半球(欧亚大陆及北美)温带地区,个别可到达南温带,但其分布中心在北温带的属。该成分的属有45个,共104种,占扣除广布成分后总属数(145属)的31.03%,占扣除广布成分后总种数(320种)的32.50%。其中,所具物种5种及以上的属有3个:丝盖伞属(10种)、乳菇属(21种)和多汁乳菇属(5种);含2-4种的属有15个:杯冠瑚菌属Artomyces (2种)、斜盖伞属Clitopilus (3种)、小杯伞属Clitocybula (2种)、蜡伞属Hygrophorus (3种)、黏滑菇属Hebeloma (4种)、岐盖伞属Insperma (2种)、鳞伞属Pholiota (3种)、网柄牛肝菌属Retiboletus (3种)、黏柄小菇属Roridomyces (2种)、乳牛肝菌属Suillus (3种)、小乳牛肝菌属Suillellus (2种)、肉杯菌属Sarcoscypha (4种)、口蘑属Tricholoma (3种)、粉孢牛肝菌属Tylopilus (3种)和干蘑属Xerula (2种)。具有2种及以上北温带成分的属有18个,共有77种,占扣除广布成分后总属数(145属)的12.41%,占扣除广布成分后总种数(320种)的24.06%,这些属为广西喀斯特林区大型真菌北温带成分的主要组成属,且主要分布于本研究中位于桂西北的雅长保护区和木论保护区。
该成分中含1个种的属有27个:网孢盘菌属Aleuria、地花孔菌属Albatrellus、类小薄孔菌属Antella、黑管菌属Bjerkandera、冬菇属Flammulina、寄生菇属Asterophora、松苞菇属Catathelasma、钉菇属色Chroogomphus、锤舌菌属Leotia、雅薄伞属Delicatula、钉菇属Gomphus、铆钉菇属Gomphidius、马鞍菌属Helvella、亚齿菌属Hydnellum、皇牛肝菌属Imperator、耙齿菌属Irpex、小疣柄牛肝菌属Leccinellum、离褶伞属Lyophyllum、小口盘菌属Microstoma、脐菇属Omphalia、栓齿菌属Phellodon、波斯特孔菌属Postia、小肉齿菌属Sarcodontia、暗盘菌属Plectania、陀螺菌属Turbinellus、核瑚菌属Typhula和玉成孔菌属Yuchengia。这些属占扣除广布成分后总属数(145属)的18.62%,占扣除广布成分后总种数(320种)的8.43%。这些属中,除了黑管菌属和脐菇属Omphalia在热带北缘的弄岗保护区也有分布外,其余属仅见于位于桂西北的保护区,尤其是雅长保护区。
(Ⅹ) 东亚-北美洲成分的属
指间断分布于东亚和北美洲温带及亚热带地区的属。该成分的属有4个,均只具1种,占扣除广布成分后总属数(145属)的2.76%,占扣除广布成分后总种数(320种)的1.25%。这4个属为大金钱菌属Megacollybia、兰茂牛肝菌属Lanmaoa、隐孔菌属Cryptoporus和多根地花菌属Polypus。
(ⅩⅠ) 东亚成分的属
指主要分布于东亚(中国、朝鲜、韩国、日本及俄罗斯远东地区)的属种,它们有时常向南延伸至我国南部甚至中南半岛,向西可达印度、尼泊尔乃至巴基斯坦。该成分的属有2个,共有3种,占扣除广布成分后总属数(145属)的1.38%,占扣除广布成分后总种数(320种)的0.94%。这2个属为棒束孢属Isaria (2种)和红皮孔菌属Pyrrhoderma (1种)。
(ⅩⅡ) 中国特有成分的属
指目前仅知道在我国有分布且向南有时也见于中南半岛的属。该成分的属有5个,均只具1种,占扣除广布成分后总属数(145属)的3.45%,占扣除广布成分后总种数(320种)的1.56%。这5个属为:裘氏牛肝菌属Chiua、黑牛肝菌属Nigroboletus、假笼头菌属Pseudoclathrus、拟异薄孔菌属Neodatronia和绒柄多孔菌属Tomenstipella。其中,绒柄多孔菌属为本研究拟定的新属,仅见于弄岗保护区和木论保护区。
通过前面对泛热带成分和北温带成分属的分析,可以看出位于热带北缘的弄岗保护区的真菌区系与位于桂西北的雅长保护区和木论保护区的大型真菌区系明显不同,因而有必要对它们进行单独的分析。
表8分别对处于广西不同纬度带的3个喀斯特地貌国家级自然保护区大型真菌属的区系成分进行了分析。结果表明,这3个保护区的泛热带成分属数占非广布成分总属数的比率依次为弄岗保护区(80.33%)>木论保护区(60.81%)>雅长保护区(50.89%),均大于50%,说明这3个保护区的大型真菌区系表现出较强的泛热带成分,且随着纬度的增加,泛热带成分的比率递减。相反,北温带成分的属比率则依次增加:雅长保护区(33.04%)>木论保护区(24.32%)>弄岗保护区(4.92%)。雅长保护区和木论保护区均处于桂西北地区,因而具有相近的属区系成分比率。雅长保护区的海拔相对较高,因而属的北温带成分较木论保护区丰富。
表8 广西不同纬度带3个喀斯特地貌保护区大型真菌属的分布区类型统计
Table 8
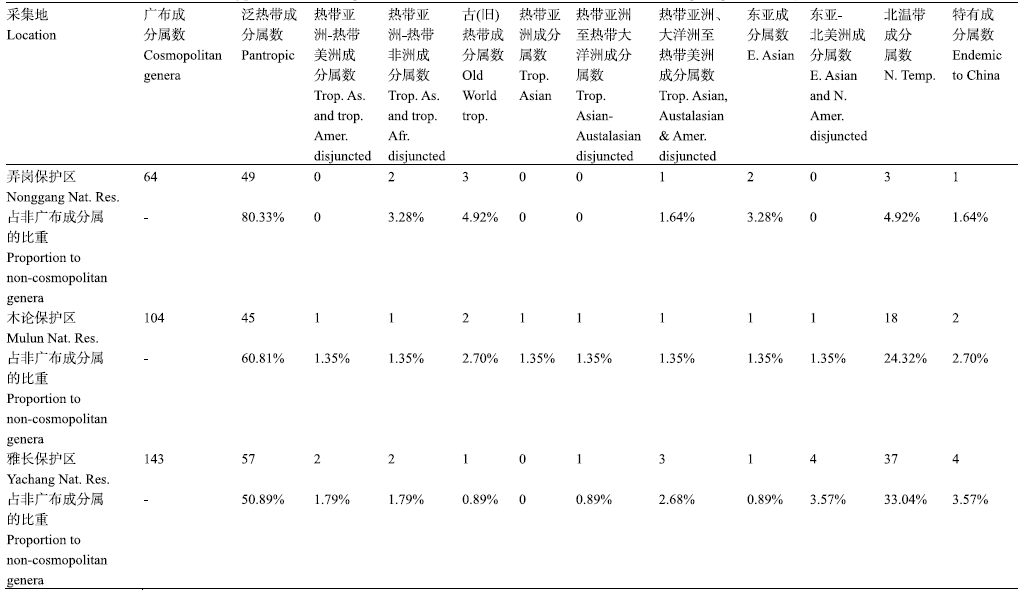 |
(3) 优势属
前面已经进行了属的组成及结构分析,本研究将属内种数在5种及以上的属以及部分属内种数小于5种,但属内物种在广西喀斯特林区十分常见的属确定为广西喀斯特林区大型真菌的优势属。
其中,子囊菌门优势属有6个(40种):毛杯菌属、炭团菌属、线虫草属、歪盘菌属、肉杯菌属和炭角菌属。这些属占子囊菌门总属数(31属)的19.35%、总种数(74种)的54.05%。
担子菌门优势属有37个(300种):蘑菇属、鹅膏属、木耳属、毛筐菌属、小鬼伞属、拟鬼伞属、丝膜菌属、靴耳属、黄囊菇属、棱孔菌属、灵芝属、老伞属、裸脚伞属、锈革菌属、丝盖伞属、乳菇属、多汁乳菇属、香菇属、白环蘑属、马勃属、红菇属、小皮伞属、微皮伞属、小孔菌属、小蘑菇属、小菇属、黑斑根孔菌属、光柄菇属、柄杯菌属、多孔菌属、小脆柄菇属、枝瑚菌属、绒盖伞属、蚁巢伞属、革菌属、栓孔菌属和银耳属Tremella。这些属占担子菌门总属数(287属)的12.89%、总种数(703种)的42.67%。
(4) 表征属
子囊菌门表征属有4个(31种):毛杯菌属Cookeina、歪盘菌属Phillipsia、肉杯菌属Sarcoscypha和炭角菌属Xylaria。这些属占子囊菌门非广布成分总属数(14属)的28.57%、总种数(45种)的68.89%。
担子菌门表征属有26个(128种):毛筐菌属、青褶伞属、鳞盖伞属、棘刚毛菌属、胶孔菌属、棱孔菌属、灵芝属、老伞属、蜂窝孔菌属、锈革菌属、丝盖伞属、乳菇属、多汁乳菇属、香菇属、白鬼伞属、小皮伞属、小蘑菇属、新大孔菌属、小奥德蘑属、柄杯菌属、须瑚菌属、黏柄小菇属、辛格杯伞属、蚁巢伞属、四角孢属和热带孔菌属。这些属占担子菌门非广布成分总属数(130属)的20%、总种数(275种)的46.55%。
3 讨论
本研究对大部分疑难标本进行了ITS序列的测序和比对,并结合形态学特征比较,确认了它们属的分类地位,直至鉴定到物种。然而,由于部分序列比对的相似度都较低,加上有些未定种大多数是大型真菌形态分类研究中比较难的类群或研究得比较少的类群,因而暂时无法定种。实际上,国内其他地方报道的区系名录中,也都有一定比例的未定种标本,这可以说是当前大型真菌研究中不可避免的问题,尤其是对于多样性丰富但调查薄弱的广西喀斯特地区。属作为一个比较自然的分类单元,在区系分析中也能够较好地反映一个地区的区系特点,尤其对于物种多样性还不是很清楚、地区间区系调查不均衡、物种分类鉴定相对困难的大型真菌来说,属在区系分析中的可靠性可能比种还高。这些已定属但未定种的标本的加入,可以提高属水平区系分析的可靠性,更能准确反映一个地区大型真菌区系的特点。这个处理方式,可能是未来很长一段时间内,对大型真菌区系研究来说,是一个相对恰当、相对准确地反映一个地区区系情况的办法。
区系调查不够充分,可以说是当前大型真菌区系地理学研究面临的最大问题之一。将基于metabarcoding技术的大型真菌物种探查与基于大型真菌子实体的区系物种调查相结合进行大型真菌区系研究,将有助于解决区系调查不充分的问题。Metabarcoding技术可以通过通用引物对环境样品中真菌总DNA的一段具有鉴定信息的基因片段进行大规模扩增,然后采用高通量测序技术进行大规模快速测序,得到可操作分类单元(operational taxonomic units, OTUs),最后通过与具有可靠物种分类信息的DNA条形码数据库中的参考序列进行比对,实现物种的鉴定(曹云等 2016),从而补充因未采集到子实体而遗漏的物种。
重视并加大对大型真菌分类人才的培养力度,增加相关研究经费的投入,才能从根本上解决大型真菌区系地理学这门学科的发展问题。大型真菌区系调查和研究依赖于大量的专业人才。反观广西已有的大型真菌区系调查资料,由广西本地学者主导的调查可以说是屈指可数。大型真菌区系问题,同时也是多样性问题。桂西南喀斯特地区作为我国生物多样性保护优先区,然而该地区大型真菌多样性情况尚不清楚,相关多样性的保护和评估工作仍然缺乏依据。只有加快相关学科的布局和建设,加强对广西本土真菌分类人才的培养,加大对广西大型真菌多样性研究的支持力度,才能尽快摸清广西大型真菌的区系情况,才能为生物多样性的保护与评估提供相对缺乏的亚热带地区大型真菌多样性数据,才能为大型真菌资源的合理开发利用提供便利。
致谢
感谢广西壮族自治区中国科学院广西植物研究所、木论国家级自然保护区管理中心、弄岗国家级自然保护区管理中心和雅长国家级自然保护区管理中心在调查采集过程中的支持和帮助。
参考文献
Fungal diversity in Daqinggou Nature Reserve
Study on fungal flora diversity in Daqinggou Nature Reserve
A monograph of Mycenaceae (Agaricales) in China
Flora fungorum sinicorum (Vol. 49 Strophariaceae Ⅰ)
Historical biogeography and diversification of truffles in the Tuberaceae and their newly identified southern hemisphere sister lineage
DOI:10.1371/journal.pone.0052765 URL [本文引用: 1]
Multi-locus phylogeny of lethal amanitas implications for species diversity and historical biogeography
DOI:10.1186/1471-2148-14-143 [本文引用: 1]
Application of metabarcoding technology in studies of fungal diversity
Fungal diversity is threatened by climate change, land-use change, and environmental pollution, and requires urgent conservation action. Construction of the fungal species database is an important prerequisite for the study and conservation of fungal diversity. Recently developed DNA barcoding and metabarcoding technologies can provide accurate, rapid, and highly efficient identification on a large scale, and to a large extent compensate for the defects of traditional identification methods. In this paper, we review the application of metabarcoding in fungal species diversity assessment, the study of mechanisms underlying fungal diversity, and the reconstruction of fungal palaeoecology. We emphasize that the application of metabarcoding technology in fungal diversity studies is still in the primary phase, and greater efforts are needed in the construction of reliable reference databases, the optimization of experimental procedures, and updates of bioinformatics tools. Hence, we suggest enhancing cooperation among fungal taxonomists, ecologists, and computer technicians. They should work together to address problems in fungal diversity studies via metabarcoding, which would provide more sound scientific evidence for fungal diversity conservation on a large scale.
Phylogeny, divergence time estimation, and biogeography of the genus Heterobasidion (Basidiomycota, Russulales)
DOI:10.1007/s13225-014-0317-2 URL [本文引用: 1]
Species diversity, taxonomy and phylogeny of Polyporaceae (Basidiomycota) in China
DOI:10.1007/s13225-019-00427-4 [本文引用: 1]
Hymenochaetaceae (Basidiomycota) in China
DOI:10.1007/s13225-010-0066-9 URL [本文引用: 1]
Diversity and systematics of the important macrofungi in Chinese forests
Flora fungorum sinicorum (Vol. 54 Lycoperdaceae Mycenastraceae)
Multilocus phylogenetic analyses reveal unexpected abundant diversity and significant disjunct distribution pattern of the hedgehog mushrooms (Hydnum L.)
DOI:10.1038/srep25586
[本文引用: 1]

Hydnum is a fungal genus proposed by Linnaeus in the early time of modern taxonomy. It contains several ectomycorrhizal species which are commonly consumed worldwide. However, Hydnum is one of the most understudied fungal genera, especially from a molecular phylogenetic view. In this study, we extensively gathered specimens of Hydnum from Asia, Europe, America and Australasia, and analyzed them by using sequences of four gene fragments (ITS, nrLSU, tef1α and rpb1). Our phylogenetic analyses recognized at least 31 phylogenetic species within Hydnum, 15 of which were reported for the first time. Most Australasian species were recognized as strongly divergent old relics, but recent migration between Australasia and the Northern Hemisphere was also detected. Within the Northern Hemisphere, frequent historical biota exchanges between the Old World and the New World via both the North Atlantic Land Bridge and the Bering Land Bridge could be elucidated. Our study also revealed that most Hydnum species found in subalpine areas of the Hengduan Mountains in southwestern China occur in northeastern/northern China and Europe, indicating that the composition of the mycobiota in the Hengduan Mountains reigion is more complicated than what we have known before.
Descriptions of six new species of Crepidotus from China
Flora fungorum sinicorum Vol. 59. Xylaria
Research progress on the biogeography of ectomycorrhizal fungi
Fungal diversity revisited: 2.2 to 3.8 million species
Tropic origins, a dispersal model for saprotrophic mushrooms in Agaricus section Minores with descriptions of sixteen new species
DOI:10.1038/s41598-017-05203-5
[本文引用: 1]
Agaricus section Minores contains the richest species diversity within the genus. Its Phylogeny is firstly presented by a Maximum Likelihood tree generated through DNA sequences from four gene regions of 91 species. Furthermore, a molecular dating analysis is conducted used those sequences, and it provided the divergence times of the clades within section Minores. Study showed section Minores has a tropical origin. Four main dispersal routes are proposed: (1) species from South Asia migrated through the Tibetan Plateau and reached Europe ca. 9–13 Ma; (2) species from out of South Asia dispersed to Europe in the earlier time of ca. 22 Ma; (3) species from South Asia dispersed through North Asia to Alaska, and reached West America around ca. 9 Ma; and (4) species from South Asia dispersed south and reached Oceania by at least three invading events about ca. 9, 12 and 16–18 Ma respectively. Those routes excepting the second route coincide with those of ectomycorrhizal mushrooms. To know whether the second route existed in the saprotrophic mushrooms requires further studies, and the fourth route may explain why the secotioid species occurring in Australia are morphologically similar but cluster in different phylogenetic clades. This study also demonstrates a great biodiversity of A. section Minores in China. Sixteen new species and three new records are introduced from China with morphological descriptions, illustrations, color photographs and phylogenetic analyses.
Shiitake mushrooms and molecular clocks historical biogeography of Lentinula
DOI:10.1046/j.1365-2699.2001.00528.x URL [本文引用: 1]
Delimitation of Funga as a valid term for the diversity of fungal communities: the Fauna
Research progress and prospect on biogeography of birds in China
Progress and perspectives on insect biogeography in China
Insects are the most diverse group of organisms on the Earth. There are more than one million insect species, accounting for more than half of the total. Insects usually play important parts in the ecosystem and have close relations with humans. Therefore understanding biodiversity and distribution patterns of insects is of important signficance for biogeography science as well as the development of human society. Based on detailed literature retrieval and content analysis, this paper presents the trends of publications on insect biogeography in China during the past 70 years, and summarizes main research progress from the following aspects: insect species survey, insect fauna, insect community diversity, genetic diversity patterns of insects, and distribution patterns of insects. Encouraging progress on insect biogeography have been made in China during the past decades, however, researchers need to strengthen the thinking and study on several aspects including integrative biogeography thinking, spatial and temporal scales, refining scientific questions and hypotheses, trait biogeography of insects, and sharing of biodiversity and distribution data of insects.
Flora fungorum sinicorum. Vol. 45 Pleurotoid-Lentinoid fungi
Atlas of Chinese macrofungal resources
Flora fungorum sinicorum. Vol. 23. Sclerodermatales Tulostomatales Phallales Podaxales
Phylogeography and biogeography of fungi
DOI:10.1016/j.mycres.2008.02.002
PMID:18346884
[本文引用: 1]
The rigorous study of processes shaping geographic distributions of lineages is a relatively new and emerging field in mycology. While it was previously generally believed that most fungi have wide distributions and largely unstructured populations, recent studies have shown that this is not the case. The study of distributions in tandem with molecular approaches to phylogeny has recently made substantial advances to our understanding of the diversity and biogeography of fungi. Comprehensive species inventories have provided a better picture of the actual distribution of these organisms, while robust phylogenies based on molecular characters have provided both data that allow interpretation of current distributions and testable hypotheses regarding the processes responsible for distribution patterns. This commentary provides an introduction to five papers in this issue of Mycological Research that focus on fungal phylogeography. These papers are based on oral contributions given at two symposia at the International Mycological Congress (IMC8) held in Cairns (Australia) in August 2006.
Out of the Palaeotropics? Historical biogeography and diversification of the cosmopolitan ectomycorrhizal mushroom family Inocybaceae
DOI:10.1111/jbi.2009.36.issue-4 URL [本文引用: 1]
Funga and resource, a case in karst forest of Guangxi
Asproinocybaceae fam. nov. (Agaricales, Agaricomycetes) for accommodating the genera Asproinocybe and Tricholosporum, and description of Asproinocybe sinensis and Tricholosporum guangxiense sp. nov
DOI:10.3390/jof7121086
URL
[本文引用: 1]
Asproinocybe and Tricholosporum are not well known, and their placement at the family level remains undetermined. In this study, we conducted molecular phylogenetic analyses based on nuc rDNA internal transcribed spacer region (ITS) and nuc 28S rDNA (nrLSU), and a dataset with six molecular markers (ITS, LSU, RNA polymerase II largest subunit (RPB1), RNA polymerase II second largest subunit (RPB2), 18S nuclear small subunit ribosomal DNA (nrSSU), and translation elongation factor 1-alpha (TEF1-α)) using Bayesian (BA) and Maximum Likelihood (ML) methods, we found that the species of Asproinocybe and Tricholosporum formed an independent family-level clade (0.98/72). Asproinocybaceae fam. nov., a new family, is established here for accommodating this clade. Two new species, Asproinocybe sinensis and Tricholosporum guangxiense, from subtropical and tropical karst areas of China, are also described here.
Plectania lutea (Sarcosomataceae), a new species from southwestern karst areas of China
Report on comprehensive survey of Longgang Reserve, Guangxi
Considering sampling methods for macrofungi exploration in Turgo tropical forest ecosystem
DOI:10.22146/jtbb.38381
URL
[本文引用: 1]
Diversity of macrofungi is widely studied because of its role as decomposer, and highly reported of its potentially using for foods, agricultures, medicinal purposes, and bioremediation. This study was carried out to consider sampling methods for macrofungi exploration especially in Turgo tropical forest ecosystem. There were two sampling method models which adequately bring our macrofungi collection to have valid data. The first is adaptive sampling which includes fixed-size plots where prior we decided how many tracks, plots, or what size to examine at each site to help keeping our focus on track. By having a fixed plot, we allowed ourselves to find our macrofungi and work on the identification process thoroughly without racing against the time as much. Then we can obtain frequency, distribution, density and others ecological quantitative data by sampling plots. Another way, we can choose opportunistic sampling, we mean carefully walking on through chosen site as study area and collecting conspicuous sporocarp of macrofungi. This method needs no plots and does not produce any quantitative data but looking for many more diversity on macrofungi. The way of sampling, depends on the duration of observation and the purposes of the research. According to the consideration result, we concluded to choose the adaptive sampling method to explore the macrofungi in Turgo tropical forest ecosystem.
Species diversity, distribution patterns, and substrate specificity of Strobilurus
DOI:10.1080/00275514.2018.1463064 URL [本文引用: 1]
A biogeographical overview of the Canadian mushroom flora
DOI:10.1139/b89-384
URL
[本文引用: 1]
Maps showing the North American distributions of 74 species of fleshy fungi, mainly Agaricales, are used to demonstrate variations in fungal ranges. Each map is based on examined specimens or selected literature. The maps are arranged to show North American floristic patterns, while the species in the text are grouped along worldwide patterns. Range patterns resemble those reported for vascular plants, ascolichens, and bryophytes. The names Hypholoma flavifolium (Smith) comb.nov. and Strobilurus trullisatus var. montezumae (Singer) comb. & stat.nov. are proposed.
Evolutionary history of Serpulaceae (Basidiomycota) molecular phylogeny, historical biogeography and evidence for a single transition of nutritional mode
Preliminary analysis of macrofungal flora of Dinghushan Biosphere Reserve in China
The macrofungal diversity in the Nanling Mountains
DOI:10.13284/j.cnki.rddl.003053
The Nanling Mountain region of South China is very rich in macrofungal resources, and also is the largest oasis near 25 degrees north latitude. The macrofungal diversity in the Nanling Mountains was introduced in this paper. 37 species originally described from the type localities in the Nanling were listed, including the Latin name, Chinese name, habitat and distribution; some misidentification or questionable species were annotated; the dominant families or genera, floristic elements, resource protection and sustainable utilization in the Nanling Mountains were analyzed. Strengthening the researches and protection of macrofungal diversity in the Nanling Mountains is greatly important not only for scientific significance, but also for ecosystem stability, regional environmental safety and future industrial development.
Current research and development trends in floristic geography
This paper summarizes the research status, existing issues, and trends in floristic geography. There is now a wealth of research accumulation on floristic investigations, distribution types of genera, floristic regions, and regional floristic analysis. It is also noted that most of these studies utilize simple statistical analyses, comparative studies, traditional methods, and single subjects, to provide a basic understanding and description of the floristic phenomenon, which is lacking spatial pattern formation processes and detailed exploration of formation mechanisms. Additionally, there are still some weak and non-existent areas of botanical investigation. Many existing specimens lack detailed or accurate information and the precise identification of plant species also needs to be much improved. At the same time, when analyzing the development trends of floristic geography, with the rapid development of related disciplines, including the development of biogeography and analysis methods and improvements, floristic geography research is an area of multidisciplinary integration, comprehensive research, and analysis. On the one hand, using database information, and combining ecology, paleobotany, and geology can allow us to probe into spatial pattern formation. On the other hand, combining phylogenetics, the tree of life, and molecular biogeography allow us to reveal floristic origins and evolution. The rapid development of various disciplines has given rise to a large amount of data, meanwhile, the emergence and application of new analytical methods and theories incorporate big data into floristic geography research, which will enhance qualitative understanding and description, and allow us to further explore the mechanisms of formation quantitatively.
Overview of nomenclature novelties of fungi in the world and China (2020)
<p id="p00005"><strong>Aims:</strong>Fungi have the richest biodiversity on earth, second to insects. Thousands of new fungal species have been discovered and described every year recently. Timely analysis of the published data of new fungal names can show the research hotspots of fungal taxonomy in details and provide reference data for the future development of mycology.</p> <p id="p00010"><strong>Method:</strong> The taxonomic data of new published fungal name were retrieved from the database of Fungal Names, Index Fungorum and MycoBank.</p> <p id="p00015"><strong>Results:</strong> In 2020, at least 4,996 new fungal names including 652 new higher taxa, 2,905 new species and new intraspecific taxa, 1,342 new combinations, and 97 other novelties, were published by 1,978 scholars in 885 articles and 9 monographs all over the world, reaching the historically highest annual record of new fungal names. These new names belong to 12 phyla, 44 classes, 173 orders, 469 families and 1,386 genera, among which ascomycetes had received more attentions. The type specimens of new species were from 103 countries and regions in the world. East and Southeast Asia were the hot spots of new species discovery, while China was the country with the highest number of new species discovered (669 species), accounting for 23% of the world's total. Besides, China had made remarkable achievements in fungal taxonomy in 2020, with 333 Chinese scholars participating in the publications of 1,271 new fungal names, accounting for about a quarter of the total number in the world.</p> <p id="p00020"><strong>Conclusion:</strong> As shown by the analysis, the discovery of new fungal species is accelerating, with the increased number of new published names and authors. However, the description rate of fungal species is still at a low level, which leaves a long-term and arduous task for mycologists to discover undescribed fungi efficiently.</p>
Species diversity and floristic composition of polypores in the southern parts of Hengduan Mountains, Southwest China
A comparison of polypore funga and species composition in forest ecosystems of China, North America, and Europe
Comparison of polypore florae and diversity from temperate to subtropical forest zones in China
<p id="p00010"><strong>Background:</strong> Polypores are an important group of wood-decaying fungi and have important ecological functions and economic values. The unique geographical structure and complex vegetation types in China provide abundant substrates for the growth of polypores. Although the taxonomy of polypores has been well studied in China, few comparative studies on the diversity and flora of polypores among different climatic zones has been performed. This study is intended to compare the florae and populations of polypores in multiple forest zones with different climates and vegetations, and will provide the basis for the protection of species diversity and the development of application potential of polypores. <br><strong> Methods:</strong> In this study, we comparatively analyzed the polypore species, ecological habits and floral characteristics in the Altai Mountains, the Qinling Mountains and the Nanling Mountains which are respectively located in boreal, temperate and subtropical zones in China. <br><strong> Results:</strong> A total of 287 poroid wood-decaying fungal species belonging to 107 genera, 29 families, and 8 orders were obtained from the three mountain ranges. Of these species, 84, 132 and 160 were found in the Altai Mountains, the Qinling Mountains and the Nanling Mountains, respectively, and the community was dominated by families Polyporaceae and Hymenochaetaceae. In the generic and species levels, 25 genera and 14 species were common in the three mountain ranges. Floristic analysis found that polypores in the Altai Mountains and the Qinling Mountains were dominated by the cosmopolitan and the northern temperate elements, while polypores in the Nanling Mountains were dominated by the cosmopolitan and the pantropical taxa. In terms of host preference, polypores in the Altai Mountains preferentially grow on gymnosperms over angiosperms, while the opposite was true in the Qinling Mountains and Nanling Mountains. In the two rotting types caused by polypores, the species number of white rot polypores increased, but that of the brown rot decreased, gradually from boreal to subtropical zones. <br><strong> Conclusion:</strong> Climate and vegetation types are the major influencing factors on the flora composition of polypores, based on the analysis of species diversity, host preference and decaying type of polypores in the three mountains.</p>
Discussion on Chinese floristics
Floristics of seed plants from China
Atlas of the Chinese species of Amanitaceae
Flora fungorum sinicorum. Vol. 52 Fungi Lepiotoidei (Agaricaceae)
Tropical Affinities of Higher Fungi in Southern China
Wood-inhabiting fungi in southern China. 6. Guangxi Autonomous Region
DOI:10.5735/085.049.0605 URL [本文引用: 1]
Floral composition and ecology of polypores in Hainan Province
The floral composition and ecological habits of polypores from Hainan Province were evaluated. Polypores were very rich in the province, and 242 species belonging to 7 orders, 17 families and 84 genera were found. The dominant families were the Polyporaceae, Hymenochaetaceae and Ganodermataceae, the dominant genera were Phellinus, Ganoderma and Polyporus, and 28, 14 and 13 species were respectively found in these genera. The majority of the polypores in Hainan were the pantropical elements and the northern temperate elements, showing distinct pantropical characteristics of floral composition. Among the 242 polypores, 102, 95 and 45 species were respectively considered as the common, occasional and rare species; 225 species were found on angiosperm wood and 38 species live on gymnosperm wood, while 21 species occur on both angiosperm and gymnosperm wood. Ecologically, these polypores can be divided into three types: Saprophytic (218 species), parasitic (20 species) and symbiotic (4 species); 225 species cause white rot and 17 species cause brown rot.
The genus Retiboletus in China
DOI:10.3852/15-072 URL [本文引用: 1]
The floristic phytogeography of spermatophyte flora in Luoxiao Range
Introductory about karst forest of Mulun Nature Reserve
Chinese polypore diversities: species, mycota and ecological functions
DOI:10.3724/SP.J.1003.2013.08074
Polypores are the group of macro-basidiomycetes with poroid hymenophore and corky basidiocarps that mainly grow on wood. China contains multiple climatic zones and geographic topographies, and thus possesses a variety of forest and vegetation types that provide rich habitats for polypores. Based on extensive collections conducted over a long period of time, a considerable knowledge of Chinese polypore species has been obtained, and this has been enriched further in recent years by using molecular technology. China possesses the highest polypore diversity in the world and 704 polypore species, belonging to 134 genera, 22 families and 11 orders, have been recorded within its borders. These 704 species are composed of cosmopolitan, boreal/temperate and tropical-subtropical elements. White-rot polypores are considered as potential industrial resources, while brown-rot polypores play an essential role in forest renewal.
Preliminary studies on species diversity of wood-decaying fungi from Guangxi Autonomous Region
Species diversity, phylogeny, divergence time, and biogeography of the genus Sanghuangporus (Basidiomycota)
DOI:10.3389/fmicb.2019.00812 URL [本文引用: 2]
Flora fungorum sinicorum. Vol. 21 Hyaloscyphaceae, Sarcoscyphaceae et Sarcosomataceae
Metabarcoding技术在真菌多样性研究中的应用
DOI:10.17520/biods.2016096
[本文引用: 1]
由于受到气候变化、土地利用变化及环境污染等诸多因素的干扰, 真菌多样性受到不容忽视的威胁, 亟需得到保护。构建物种数据库是实现真菌多样性研究和保护的重要前提。近年来兴起的DNA条形码及metabarcoding技术能够在很大程度上弥补传统鉴定方法的缺陷, 可对真菌物种进行大规模、准确、快速、高效地鉴定。本文梳理了metabarcoding技术在真菌物种多样性评估、真菌多样性影响机制和真菌古生态重建等研究中的应用, 同时强调了metabarcoding技术用于真菌多样性研究尚处于初期阶段, 在构建有效参照数据库、优化实验流程以及升级生物信息学工具等方面仍需要进一步的完善。建议加强真菌分类学家、生态学家以及计算机工具研发工程师之间的合作, 共同解决metabarcoding技术在真菌多样性研究及应用中面临的问题, 为宏观尺度上真菌多样性保护提供更加科学的依据。
中国森林大型真菌重要类群多样性和系统学研究
中国昆虫生物地理学进展与展望
DOI:10.11821/dlxb202201010
[本文引用: 1]
昆虫是地球上多样性最高的生物类群,其物种数量超过所有生物物种数量的一半,在生态系统中具有重要功能,且与人类生活密切相关。理解昆虫多样性及地理分布格局对于科学研究和人类社会发展有重要意义。基于详细的文章资料梳理,本文总结了1950—2020年中国昆虫生物地理学领域文章发表趋势,并从昆虫物种多样性调查、昆虫区系研究、昆虫群落多样性、昆虫遗传多样性格局、昆虫地理分布格局等几个方面论述了中国昆虫生物地理研究的代表性研究进展。中国昆虫生物地理学研究几十年来取得了可喜的发展,但仍需重点加强几方面的思考和工作,包括整合性思维、时空尺度、科学问题的凝练、昆虫性状生物地理学、昆虫多样性和地理分布数据共享。
广西喀斯特林区大型真菌区系地理学与资源学研究
植物区系地理研究现状及发展趋势
DOI:10.17520/biods.2016253
[本文引用: 1]
中国植物区系地理学在植物区系的调查、分布区类型的划分和研究、植物区系分区以及区域植物区系的分析等方面有了丰富的研究积累。但这些研究绝大部分是利用传统方法对植物物种或分类群数据进行统计分析, 因此仍处于对区系现象的认识和描述阶段, 缺乏空间格局形成过程、形成机制方面的深入探索。此外, 还存在植物区系调查薄弱甚至是空白的地区, 许多植物标本缺乏详细或精确的信息, 植物物种鉴定的准确性还需要提高。同时, 随着植物区系地理以及相关学科的快速发展, 植物区系地理学的研究步入了多学科交叉融合、综合研究分析的阶段, 学科界限趋于模糊。一方面, 利用物种信息数据库, 融合生态学、古植物学以及地质历史等探讨区系空间地理格局的成因; 另一方面, 结合系统发育、分子系统进化(甚至到进化基因组学)、生命之树以及分子生物地理学等揭示区系的起源和进化; 同时, 各学科的快速发展, 产生了大量的数据, 新的分析方法或新理论的出现和运用, 使植物区系地理研究也进入到了大数据时代, 进而使得植物区系地理研究由定性的现象认识和描述, 逐步深入到了定量地解释区系现象和探索其形成机制的时代。
世界及中国菌物新命名发表概况(2020年)
DOI:10.17520/biods.2021202
[本文引用: 2]
菌物是全球生物多样性最丰富的生物类群之一, 每年有大量新物种被描述和发现, 统计和分析菌物新名称发表数据, 可实时展示菌物分类学的研究热点, 并为学科的未来发展提供参考数据。根据世界菌物名称信息库Fungal Names、Index Fungorum和MycoBank所收录的数据, 2020年全球共发表了4,996个菌物新名称, 包括652个新高阶分类单元、2,905个新种及种下单元、1,342个新组合和97个其他名称, 是历史上发表菌物新名称数量最多的一年。这些新名称隶属于12门44纲173目469科1,386属, 盘菌、小型子囊菌、地衣和伞菌是本年度最受关注的类群。来自全球的1,978位学者将上述菌物新名称发表在133个期刊的885篇论文和9部专著中, 其中669位作者是首次参与新名称发表, 是历年来新作者最多的一年。本年度发表的菌物新物种的模式标本来自世界103个国家和地区, 东亚和东南亚是新物种发现的最热点地区, 而中国是发现新物种最多的国家, 共发现669种, 占全球的23%, 是排名第二位的泰国的2.28倍。西南地区是我国新物种发现的热点地区, 云南、贵州、西藏三省区本年度发现的新物种数量占全国的44.80%。除了新物种来源, 中国学者在本年度的菌物分类学研究中取得了令人瞩目的成果, 333位中国学者参与发表了1,271个菌物新名称, 占全球新名称总数的1/4。这些菌物新名称隶属于8门24纲87目209科440属, 发表在45个期刊的258篇研究论文和1部专著中。通过上述研究发现, 全球菌物新物种发现的速度仍在加快, 命名作者和新名称发表数量在逐步增加, 研究类群规模在逐渐扩大。但菌物物种描述率仍处于较低水平, 持续、高效地发现菌物新物种将是菌物学者的重点研究方向之一。
我国寒温带至亚热带森林多孔菌区系和多样性比较
DOI:10.17520/biods.2021094
[本文引用: 2]
多孔菌是木材腐朽菌的重要类群, 具有重要的生态功能和经济价值。本文比较分析了我国寒温带至亚热带的阿尔泰山脉、秦岭山脉和南岭山脉的多孔菌物种、生态习性和区系特征。经调查, 在三个山脉共发现多孔菌8目29科107属287种, 其中阿尔泰山、秦岭和南岭分别为84种、132种、160种, 优势科均为多孔菌科和锈革孔菌科。三个山脉的共有属和共有种分别为25个和14个。区系地理分析发现, 阿尔泰山脉和秦岭山脉以世界广布成分和北温带成分为主, 南岭山脉以世界广布和泛热带成分为主。在寄主选择性方面, 阿尔泰山脉的多孔菌偏好生长在裸子植物上, 其比例高于被子植物, 而秦岭和南岭则相反。在腐朽类型方面, 从寒温带至亚热带白腐真菌物种数量呈现逐渐上升的趋势, 而褐腐真菌数量逐渐下降。通过比较分析3个不同气候带的多孔菌物种多样性、寄主偏好性和引起的腐朽类型, 发现气候和植被类型是影响多孔菌区系组成的主要因素。
中国海南省多孔菌区系组成和生态习性
对海南省的多孔菌区系组成和生态习性进行了分析,发现该地区多孔菌物种多样性十分丰富,共242种,隶属7目17科84属,优势科为多孔菌科、锈革孔菌科和灵芝科,优势属为木层孔菌属、灵芝属和多孔菌属,分别为28、14和13种。经过区系地理分析发现,海南省多孔菌以泛热带成分和北温带成分为主,表现出明显的泛热带特征。该地区的多孔菌常见种、偶见种和稀有种分别为102、95和45种。在寄主选择性方面,生长在被子植物上的有225种,裸子植物上38种,同时能够在被子植物和裸子植物上生长的有21种;在寄主专化性方面,218种为腐生菌,20种为寄生菌,4种共生菌;在腐朽类型方面,225种造成白色腐朽,占总种数的93%,17种造成褐色腐朽,占7%。
中国多孔菌多样性初探:物种、区系和生态功能
DOI:10.3724/SP.J.1003.2013.08074
[本文引用: 2]
多孔菌是指子实层体呈孔状且质地为革质至木质的一类大型担子菌, 主要生长在各类木材上。我国地域辽阔, 气候带和地形多样, 森林类型和植被组成丰富, 为多孔菌提供了丰富的栖息地。长期的野外资源调查和标本采集, 特别是近年来分子生物学技术的发展, 极大地丰富了对我国多孔菌种类的认识。我国是世界上多孔菌物种多样性最丰富的国家, 现在已知有多孔菌704种, 隶属于11目22科134属, 包括世界广布成分、北温带成分和热带-亚热带成分。多孔菌中的白腐菌是潜在的工业用菌, 褐腐菌在森林的更新过程中起重要作用。



{kind=link}
{kind=link}
{kind=link}
{kind=link}