



19世纪Fries (1821)成立蘑菇属帽伞族Agaricus trib. Galera Fr.,并将部分锥盖伞归入其中,并于1838年成立粪锈伞属Bolbitius Fr.,其中仅包含6个种(Fries 1838),51年后Fayod (1889)成立田头菇属Agrocybe Fayod,其中包含锥盖伞属Conocybe、粪锈伞属Bolbitius、小鳞伞属Pholiotina和脆锈伞属Naucoria (Fr.) P. Kumm.。之后,Singer (1948)成立了粪锈伞科Bolbitiaceae Singer,将田头菇属Agrocybe、粪锈伞属Bolbitius、锥盖伞属Conocybe、小鳞伞属Pholiotina和圆头伞属Descolea Singer划入其中,构建了粪锈伞科的原始框架,1975年又将小帽伞属Galerella Earle、假锥盖伞属Pseudoconocybe Hongo 2个属加到粪锈伞科Bolbitiaceae中(Singer 1975)。直到1982年 Watling (1982)将粪锈伞科分为3个属,但其将一些小鳞伞属Pholiotina物种归入锥盖伞属Conocybe中。
在20世纪70年代到至今,粪锈伞科Bolbitiaceae传统形态学分类框架是Watling和Hausknecht等人共同搭建的,Watling基于欧洲的标本材料对粪锈伞科Bolbitiaceae进行系统研究,对科内物种进行描述和框架的搭建(Watling 1971a, 1976, 1977, 1983, 1985;Watling & Bigelow 1983;Watling & Abraham 1986;Watling & Millerx 1987;Watling et al. 2011)。而1988年至1992年期间,Singer与其学生Hausknecht同样基于欧洲的标本材料进行研究,对粪锈伞科Bolbitiaceae物种补充描述,学名订正等,并为以后Hausknecht深入研究锥盖伞属Conocybe和小鳞伞属Pholiotina奠定了基础(Singer & Hausknecht 1988, 1992)。从1992年始,Hausknecht专注于锥盖伞属和小鳞伞属的研究(Hausknecht & Krisai-Greilhuber 1992),基于欧洲和北美的标本材料,于2009年出版关于锥盖伞属Conocybe和小鳞伞属Pholiotina专著(Hausknecht 2009),并发表一系列文章(Hausknecht & Enderle 1992;Hausknecht & Krisai-Greilhuber 1992, 2004, 2009;Hausknecht 1993, 1995, 1996, 1998, 1999, 2002a, 2002b, 2003, 2005;Hausknecht et al. 2007;Kalamees et al. 2013),并于2009年描述了关于亚洲的锥盖伞属Conocybe和小鳞伞属Pholiotina物种(Hausknecht & Krisai-Greilhuber 2009)。继Watling对粪锈伞科Bolbitiaceae物种的研究之后,锥盖伞属Conocybe及小鳞伞属Pholiotina的物种显著增多,成为了粪锈伞科Bolbitiaceae的核心成员,也引得更多学者参与其中,如Enderle,同Hausknecht一样,对锥盖伞属Conocybe和小鳞伞属Pholiotina进行研究,并且合作发表过关于锥盖伞属和小鳞伞属的文章(Enderle 1991a, 1991b, 1994, 1996;Hausknecht & Enderle 1992;Enderle & Hübner 1999)。
而对于锥盖伞属Conocybe与小鳞伞属Pholiotina的关系以及属下划分,不同学者的看法不尽相同,Watling (1970, 1971b)和van Waveren (1970)早期就将小鳞伞属Pholiotina认为是锥盖伞属Conocybe下的亚属,而Fayod (1889)、Singer & Hausknecht (1988)和Arnolds (2005)认为小鳞伞属Pholiotina应当单独为属。后来Hausknecht & Krisai-Greilhuber (2006, 2007, 2009)和Hausknecht & Contu (2007)的研究证实了小鳞伞属Pholiotina应单独成属,并对其属下框架进行探讨和调整。Watling et al. (2011)后期也赞同了这一观点,并在其文中也有所体现。
继分子生物学手段在系统分类中应用后,早期锥盖伞和小鳞伞属系统学研究多见于其他类群的系统学研究论文。Hopple & Vilgalys (1999)基于SSU片段,采用MP法,以粪锈伞属Bolbitius、锥盖伞属Conocybe、田头菇属Agrocybe、球盖菇属Stropharia (Fr.) Quél. 和垂幕菇属Hypholoma (Fr.) P. Kumm. 为外类群,研究鬼伞类真菌及其他暗色孢子伞菌分类系统学时发现田头菇与锥盖伞关系较远,而与球盖菇属Stropharia和垂幕菇属Hypholoma (Fr.) P. Kumm.物种关系较近,应排除在锥盖伞相关类群以外。Moncalvo et al. (2000, 2002)研究伞菌型真菌的系统发生关系发现锥盖伞属Conocybe单系,且与斑褶菇类真菌构成姊妹类群,2002年,他采用nLSU序列对877个分类群基于MP和Bayers法构建系统发育树,结果显示腹伞属Gastrocybe Watling与锥盖伞属Conocybe聚为一支,构成锥盖伞型分支Clade Conocyboid,而小鳞伞属Pholiotina与粪锈伞属Bolbitius亲缘较近构成粪锈伞型分支Clade Bolbitoid,2支为姊妹群,同时也验证了田头菇与锥盖伞及相关类群亲缘关系较远;Matheny et al. (2006)采用6基因片段以Bayers法和MP法分析伞菌目的系统发生框架,研究证实了Moncalvo的结果。Tóth et al. (2013)联合ITS、nrLSU和EF1-α基因片段构建锥盖伞属及相关属的系统发生框架,结果显示锥盖伞属单系,而小鳞伞属多系来源。
我国关于该2属的研究较少,以戴芳澜(1979)记载国内锥盖伞属开始,记载物种较多的有李玉,图力古尔和刘敬等人(谢支锡等 1986;李建宗等 1993;卯晓岚等 1993;毕志树等 1994;袁明生和孙佩琼 1995, 2007;张树庭和卯晓岚 1995;臧穆 1996;卯晓岚和庄剑云 1997;黄年来 1998;卯晓岚 1998, 2000;邵力平和项存悌 1998;图力古尔等 2001, 2007, 2014;李玉和图力古尔 2003; 吴兴亮等 2010;Li & Azbukina 2011;王薇 2014;李玉等 2015;图力古尔 2016;李保卫等 2016;王锋尖等 2017;刘敬 2018;张泽文 2019;冶晓燕 2021),共记载我国锥盖伞32种,小鳞伞15种,且多数情况下将小鳞伞物种作为锥盖伞处理,2属界限未十分明晰。目前,学者们普遍接受Hausknecht (2009)对该2属关系划分和属下等级划分的观点,但依然存在混淆现象。
该类真菌多生于肥沃土壤、食草动物粪便和植物残体,属于腐生菌(Matheny et al. 2006;Tóth et al. 2013)。锥盖伞和小鳞伞属中的物种含有光盖伞素、光盖伞辛、鬼笔毒肽和鹅膏毒肽等有毒物质,具有神经精神毒性、胃肠炎毒性和肝损伤毒性,国内共记载4种锥盖伞为有毒蘑菇(Wu et al. 2019)。但这些有毒物质具有潜在的药理活性,如光盖伞素是一种5-羟色胺2A受体的选择性激动剂,具有致幻作用,但对顽固性抑郁症、创伤后应激障碍、集束性头痛、强迫症、吸烟成瘾等神经精神疾病及心理障碍有显著疗效(Griffiths et al. 2016;Johnson et al. 2017;Paul 2021)。
基于采自东北、华北及华南地区的标本,按照Hausknecht的属下分类观点,进行属下等级划分,其中新组合1个,中国新记录种8个:锥盖伞属5种,小鳞伞属3种。
1 材料与方法
1.1 材料和仪器
研究材料主要为作者及团队成员采自各地的标本,全部存放在吉林农业大学菌物标本馆(HMJAU)。
1.2 方法
1.2.1 野外采集
采集时首先进行原生态拍照,记录标本环境、生长基物和植被等情况,子实体颜色记录采用Kornerup & Wanscher (1963)的方法。拍照后采集的担子体采用硅胶干燥法置于采集盒中或采用锡纸包裹带回用烘干器烘干(40 ℃),对干燥后的标本登记、入库保存于标本馆。
1.2.2 室内研究
参考Liu & Bau (2018)的研究方法,光学显微镜观察干燥标本的宏观形态和显微特征,记录测量各种形态结构,并描述绘图,世界分布结合文献并参考GBIF网站(
1.2.3 分子系统学研究
参考Mou & Bau (2021)提取DNA、PCR和测序的方法,并通过NCBI下载Tóth et al. (2013)构建Bolbitiaceae框架的相关ITS序列,使用在线Mafft (t
表1 粪锈伞科自测序列
Table 1
| 种名 Species | 凭证号 Specimen No. | GenBank ITS | 来源 Locality |
|---|---|---|---|
| Bolbitius titubans B. titubans Conocybe apala C. apala C. bisporigera C. brachypodii C. cylindricaspora C. cylindricaspora C. deliquescens C. deliquescens C. dunesis C. dunesis C. echinata C. fuscimarginata C. macrocephala C. macrocephala C. mesospora C. moseri C. moseri C. pilosella C. pilosella C. pulchella C. semiglobata C. sp. C. tenera C. velutipes Pholiotina coprophila Ph. cyanopus Ph. dasypus Ph. dasypus Ph. dasypus Ph. serrata Ph. serrata Ph. serrata Ph. serrata Ph. smithii Ph. utricystidiata Ph. utricystidiata Ph. vexans | HMJAU44966 HMJAU44965 HMJAU61996 HMJAU61997 HMJAU45055 HMJAU45017 HMJAU42441 HMJAU42440 HMJAU61998 HMJAU61999 HMJAU45041a HMJAU45041b HMJAU45076 HMJAU42438 HMJAU44967 HMJAU44971 HMJAU45049 HMJAU62000 HMJAU44986 HMJAU45061 HMJAU45060 HMJAU44984 HMJAU44981 HMJAU45018 HMJAU44970 HMJAU44995 HMJAU45088 HMJAU45092 HMJAU45004 HMJAU45105 HMJAU45103 HMJAU42442 HMJAU42443 HMJAU62004 HMJAU62006 HMJAU62001 HMJAU45083 HMJAU45085 HMJAU42439 | OP391672 OP391673 OP373405 OP373406 OP526418 MH141423 MG250377 MG250375 OP373403 OP373404 OP373393 OP373394 MH141418 MK412383 MH141420 MH141421 MH141419 OP373408 OP373395 OP391668 OP391669 MH141424 MH141441 MH141422 MH141417 OP526419 OP391671 OP391670 OP373400 OP373401 OP373402 MG250376 MG250378 OP538571 OP538570 OP373407 OP373398 OP373399 MG250380 | Jilin, China Jilin, China Hunan, China Anhui, China Jilin, China Jilin, China Jilin, China Jilin, China Inner Mongolia, China Inner Mongolia, China Inner Mongolia, China Inner Mongolia, China Jilin, China Jilin, China Jilin, China Jilin, China Jilin, China Hunan, China Jilin, China Jilin, China Jilin, China Jilin, China Jilin, China Jilin, China Jilin, China Jilin, China Inner Mongolia, China Jilin, China Jilin, China Jilin, China Jilin, China Jilin, China Jilin, China Jilin, China Jilin, China Jilin, China Jilin, China Jilin, China Jilin, China |
2 结果与分析
2.1 分子系统研究结果
基于ITS构建的ML和BI系统发育树(图1)可知,小脆柄菇属Psathyrella物种为外类群,在系统树的基部;而圆头伞属Descolea作为粪锈伞科Bolbitiaceae中的一员,同样存在于基部位置,支持率为(100/1),其是菌根共生菌;其他如锥盖伞属Conocybe、小鳞伞属Pholiotina和粪锈伞属Bolbitius物种为腐生菌。
图1
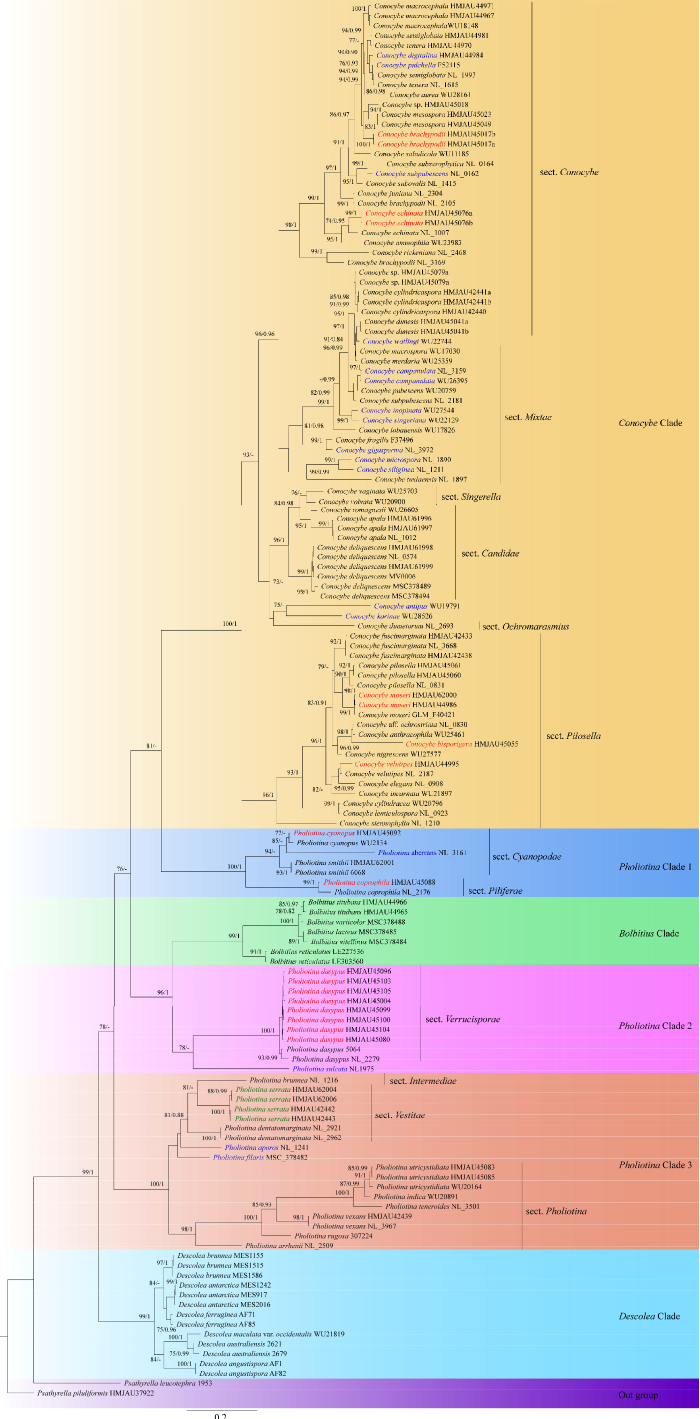
图1
基于ITS采用贝叶斯法和最大似然法构建的系统发育树
共142份标本和菌株的序列,不同颜色代表不同分支,其中蓝色字体代表其形态划分和分子不能拟合,红色字体代表为新记录种,绿色字体为新组合种
Fig.1
Phylogenetic tree constructed by Bayesian and maximum likelihood methods based on ITS sequences.
142 specimens and strains were analyzed, and different branches were distinguished by color. Among them, the blue font represents the morphological division and molecular mismatch, the red font represents the new record species, and the green font represents the new combination.
小鳞伞属Pholiotina分3个分支,分别为Pholiotina Clade 3、Pholiotina Clade 2和Pholiotina Clade 1。在Pholiotina Clade 3中,小鳞伞组sect. Pholiotina Fayod所处分支与被外菌幕组sect. Vestitae (Watling) Hauskn. & Krisai和过渡组sect. Intermediae (Watling) Hauskn. & Krisai 所处分支互为姊妹类群(100/1)。小鳞伞组sect. Pholiotina中来自吉林的近柱囊小鳞伞Ph. utricystidiata Enderle & H.-J. Hübner与来自德国的近柱囊小鳞伞Ph. utricystidiata聚在一起,支持率为(91/1);齿缘小鳞伞Ph. serrata之前被命名为齿缘锥盖伞C. serrata T. Bau & J. Liu,但在系统学的位置是处于Pholiotina Clade 3中的被外菌幕组sect. Vestitae (菌幕残留在菌盖表面),因此本研究处理为新组合。Pholiotina Clade 2与Bolbitius Clade互为姊妹类群(96/1),疣孢组sect. Verrucisporae Singer中来自吉林的糙孢小鳞伞Ph. dasypus (Romagn.) P.-A. Moreau (图2),与来自意大利和匈牙利的糙孢小鳞伞Ph. dasypus聚在一起(100/1)。而Pholiotina Clade 1则与锥盖伞分支Conocybe Clade互为姊妹类群(81/-),其中被毛组sect. Piliferae Hauskn. & Krisai和蓝色组sect. Cyanopodae (Singer) Hauskn. & Krisai同样互为姊妹类群(100/1),来自国内的材料可以和国外的材料汇聚为一支。
图2

图2
新记录种野外生境照片
A:毛柄锥盖伞. B,C:条斑锥盖伞. D:双孢锥盖伞. E:刺毛锥盖伞. F:短柄锥盖伞. G:糙孢小鳞伞. H:蓝柄小鳞伞. I:喜粪小鳞伞. 标尺=1 cm
Fig. 2
Basidiomata in situ of Chinese new record species.
A: Conocybe velutipes. B, C: Conocybe moseri. D: Conocybe bisporigera. E: Conocybe echinata. F: Conocybe brachypodii. G: Pholiotina dasypus. H: Pholiotina cyanopus. I: Pholiotina coprophila. Bars=1 cm.
Conocybe Clade则为单系类群,其在形态学分为6组:具毛组sect. Pilosellae Singer、赭小皮伞组sect. Ochromarasmius (Singer) Hauskn. & Krisai、白色组sect. Candidae Watling、小辛格组sect. Singerella Hauskn. & Krisai、混杂组sect.Mixtae Hauskn. & Krisai和锥盖伞组sect. Conocybe Fayod。其中,具毛组sect. Pilosellae为基部分支,与其余各组形成姊妹关系(100/1),且系统学结果与形态学分组一致(标红的物种为新记录种);在形态学分组中将孢子表面粗糙有疣凸的物种分到赭小皮伞组sect. Ochromarasmius,其所在的分支与白色组sect. Candidae和小辛格组sect. Singerella所在分支互为姊妹类群(73/-),白色组sect. Candidae物种是具自溶现象的分组,而小辛格组sect. Singerella的物种在国内尚未发现;在系统树进化末端的两大分支系统学结果与形态学分组不能得到很好的拟合,包含了混杂组sect. Mixtae和锥盖伞组sect. Conocybe。如此情况同样发生在其他组中(蓝色字表示的物种)。
2.2 分类
图3
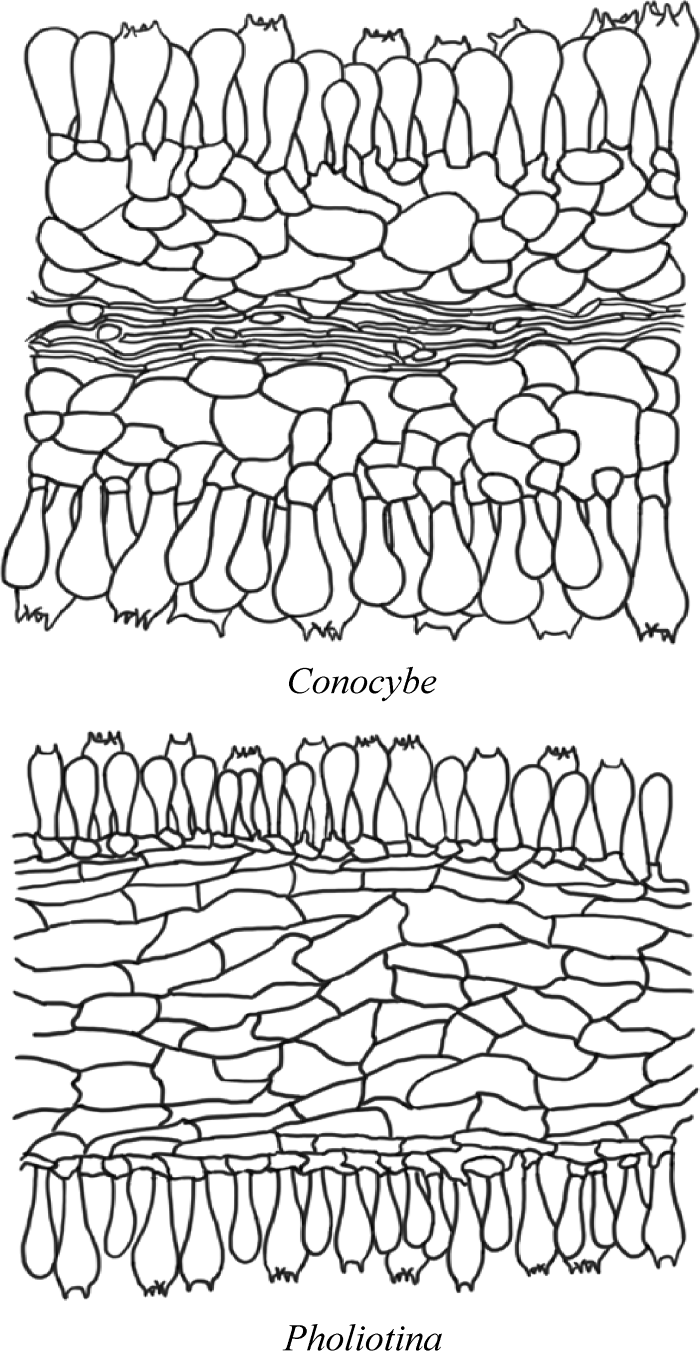
图3
Conocybe和Pholiotina物种菌褶菌髓形态
Conocybe:Conocybe semiglobata HMJAU44981 (干);Pholiotina:Pholiotina serrata HMJAU 62006 (干)
Fig. 3
Hymenophoral trama of Conocybe and Pholiotina.
Conocybe: Conocybe semiglobata HMJAU44981 (dry); Pholiotina: Pholiotina serrata HMJAU 62006 (dry).
中国锥盖伞属Conocybe和小鳞伞属Pholiotina分属检索表
1. 子实下层菌褶菌髓被压缩成窄圆柱形或近圆柱形,子实层基部菌髓气泡状;缘生囊状体全部球颈瓶形;不存在菌幕残留···········································································································锥盖伞属Conocybe
1’. 子实下层菌褶菌髓气泡状近宽柱状,菌髓中心不被压缩;缘生囊状体非球颈瓶形(泡囊状、近圆柱状、披针形和烧瓶形),若为球颈瓶形则带菌幕残留···········································································小鳞伞属Pholiotina
锥盖伞属Conocybe分组检索表
1. 担子体成熟后菌盖菌褶多自溶······························································白色组sect. Candidae
1’.担子体菌盖菌褶不自溶······················································································2
2. 孢子粗糙,具疣状点或褶皱··········································································赭小皮伞组sect. Ochromarasmius
2’. 孢子表面光滑······························································································3
3. 柄生囊状体非球颈瓶形··································································具毛组sect. Pilosellae
3’. 柄生囊状体球颈瓶形······················································································4
4. 柄生囊状体全部为球颈瓶形·························································锥盖伞组sect. Conocybe
4’. 柄生囊状体中同时存在球颈瓶形及非球颈瓶形···················································混杂组sect. Mixtae
锥盖伞属白色组Conocybe sect. Candidae分种检索表
1. 担子体成熟后菌柄基部非球茎状·········································卡氏锥盖伞C. karinae
1’. 担子体成熟后菌柄基部球茎状······································································2
2. 菌盖高宽相近,非水浸状;菌柄直··························································乳白锥盖伞C. apala
2’. 菌盖高大于宽,水浸状;菌柄常弯曲·····································融黏锥盖伞C. deliquescens
锥盖伞属具毛组Conocybe sect. Pilosellae分种检索表
1. 菌盖新鲜湿润时中心褐色··················································································2
1’. 菌盖新鲜湿润时中心无褐色··················································································6
2. 菌盖湿润时无条纹······································································石灰锥盖伞C. siliginea
2’. 菌盖湿润时具条纹······························································································3
3. 担孢子无萌发孔······································································绒毛锥盖伞C. pilosella
3’. 担孢子具萌发孔······························································································4
4. 菌柄基部无球茎 赭色锥盖伞C. ochracea
4’. 菌柄基部呈球茎状······························································································5
5. 4担孢子,明显肾形······································································毛柄锥盖伞C. velutipes
5’. 4和2担孢子混合,从不肾形·································································条斑锥盖伞C. moseri
6. 菌盖不水浸状,无条纹,略带皱纹······························································灰环锥盖伞C. fuscimarginata
6’. 菌盖水浸状,具条纹,表面光滑··················································································7
7. 菌柄基部具假根··························································肉色锥盖伞C. incarnata
7’. 菌柄基不具假根··························································································8
8. 具2个担子小梗··························································双孢锥盖伞C. bisporigera
8’. 具4个担子小梗·················································································· ········9
9. 担孢子凸镜形········································································凸镜孢锥盖伞C. lenticulospora
9’. 担孢子椭圆形········································································赭叶锥盖伞C. siennophylla
图4
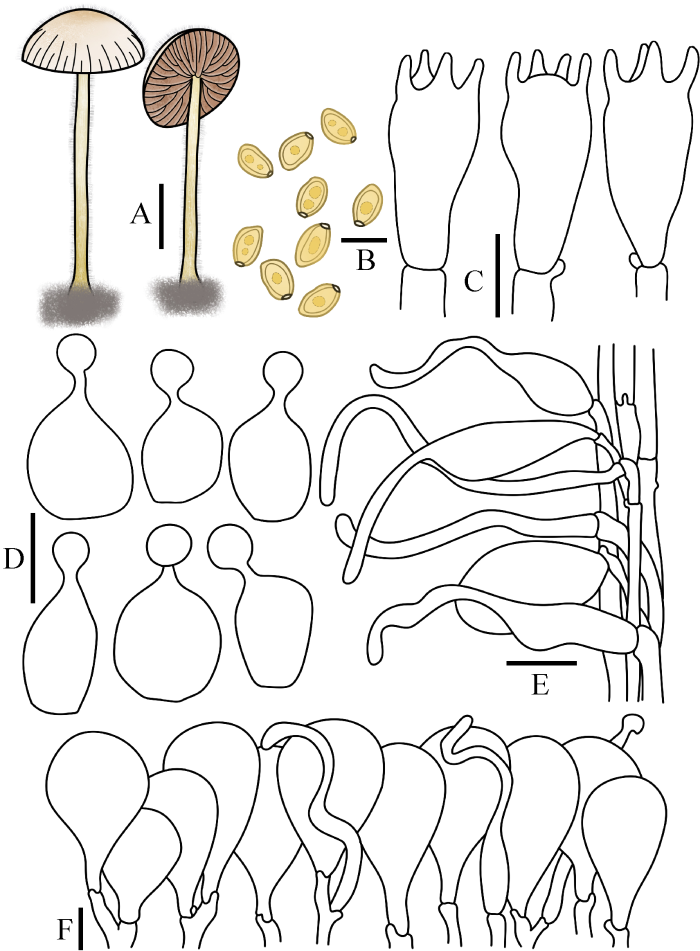
图4
毛柄锥盖伞
A:担子体. B:担孢子. C:担子. D:缘生囊状体. E:柄生囊状体. F:菌盖表皮. 标尺:A=1 cm;B-F=10 μm
Fig. 4
Conocybe velutipes.
A: Basidiomata. B: Basidiospores. C: Basidia. D: Cheilocystidia. E: Caulocystidia. F: Pileipellis. Bars: A=1 cm; B-F=10 μm.
Conocybe velutipes (Velen.) Hauskn. & Svrček, in Hausknecht, Czech Mycol. 51(1): 68 (1999)
≡ Galera velutipes Velen., Novitates Mycologicae: 128 (1939)
= Conocybe kuehneriana Singer [as 'kuhneriana'], Beih. Nova Hedwigia 29: 212 (1969)
菌盖直径0.5-2.5 cm,高可达1.0 cm,半球形至凸镜形,中心棕褐色,边缘污白色至淡黄色。菌盖表面水浸状,被白色细小绒毛,具浅沟纹。菌肉薄,污白色至淡黄色,无特殊气味。菌褶直生近弯生,稍密,略腹鼓状,幅宽2-4 mm,赭黄色至淡褐色,菌褶边缘平滑。菌柄长3.5-8.5 cm,粗1.0-2.5 mm,圆柱形,基部膨大至弱球茎状,表面被白色绒毛和粉霜,稍具纵条纹。
担孢子10.5-14×6.5-8.5 μm,Q=1.40-1.80,椭圆形至长椭圆形,稍肾形,壁厚,内含油滴,萌发孔明显,担孢子在水中呈黄色,KOH溶液中棕褐色。担子17-28×9-12 μm,棒状,具4担子小梗。缘生囊状体球颈瓶形,14-25×7-10 μm,球顶直径2-5 μm。无侧生囊状体。柄生囊状体烧瓶形,圆柱形,毛发状。菌盖菌丝膜皮型(“hymeniform”型),由24-63×12-34 μm的球茎细胞或倒梨形细胞排列而成。盖生囊状体毛发状近圆柱形。所有组织均具锁状联合。
生境:夏秋季散生于草地上。
世界分布:亚洲(中国);欧洲(瑞典、[前]捷克斯洛伐克[模式]、德国、奥地利和英国)。
中国分布:吉林、内蒙古。
研究标本:吉林:长春市吉林农业大学校园,2016年9月5日,图力古尔、刘敬,HMJAU45044;长春市吉林农业大学校园,2016年9月12日,图力古尔、刘敬,HMJAU45045;内蒙古:通辽市辽河公园,2016年8月2日,图力古尔、刘敬,HMJAU44995。
讨论:该种因其柄生囊状体为非球颈瓶形而被归入具毛组sect. Pilosellae。其主要特征为菌盖半球形至凸镜形,孢子稍肾形。该种与赭叶锥盖伞C. sienophylla (Berk. & Broome) Singer ex Chiari & Papetti菌盖形态颜色相近,但前者担孢子10.5-14×6.5-8.5 μm,后者担孢子小于前者,易于区别;该种又与雅致锥盖伞C. elegans Watling为近缘物种,显微特征相近,但后者菌盖圆锥形,因此二者易于区分(Arnolds 2005)。
图5
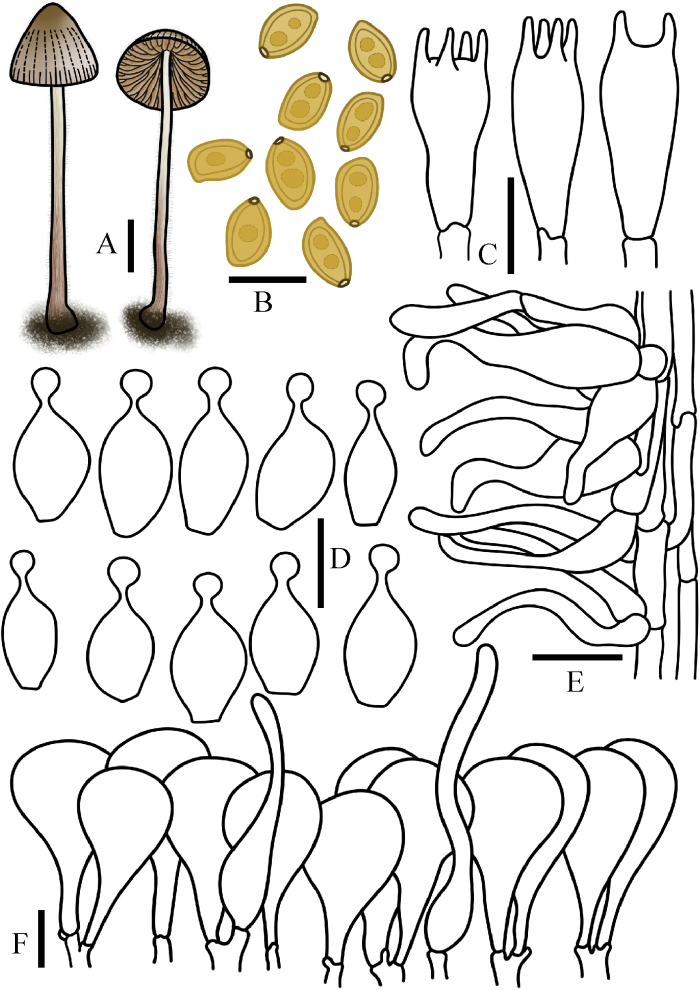
图5
条斑锥盖伞
A:担子体. B:担孢子. C:担子. D:缘生囊状体. E:柄生囊状体. F:菌盖表皮. 标尺:A=1 cm;B-F=10 μm
Fig. 5
Conocybe moseri.
A: Basidiomata. B: Basidiospores. C: Basidia. D: Cheilocystidia. E: Caulocystidia. F: Pileipellis. Bars: A=1 cm; B-F=10 μm.
Conocybe moseri Watling, Notes R. bot. Gdn Edinb. 38(2): 342 (1980)
= Conocybe kuehneri Singer, Collnea bot., Barcinone Bot. Instit. 1: 236 (1947)
= Conocybe moseri var. robustior Hauskn. & H.-J. Hübner, in Hausknecht, Öst. Z. Pilzk. 14: 244 (2005)
菌盖直径0.5-2.0 cm,钟形,圆锥形,中心灰褐色,边缘灰白色,表面具明显沟纹。菌肉稍厚,污白色至黄褐色,无特殊气味。菌褶直生近弯生,稍密,幅宽2-4 mm,黄褐色至红褐色,边缘平滑。菌柄长4.0-6.0 cm,粗1.0-1.5 mm,圆柱形,基部稍膨大,表面被白色粉霜,絮状细小绒毛,具纵沟纹。
担孢子8.5-12×5.0-7.5 μm,Q=1.33-1.82,椭圆形至长椭圆形,壁厚,光滑,内含油滴,有明显的萌发孔,担孢子在水中呈赭黄色,在KOH溶液中呈橙褐色。担子16-22×7-12 μm,棒状,具4(2)担子小梗。缘生囊状体球颈瓶形,15-21× 5-10 μm,球顶直径2-5 μm。无侧生囊状体。柄生囊状体由近圆柱形,发丝状,圆柱形组成。菌盖菌丝膜皮型(“hymeniform”型),由20-44× 10-27 μm的球茎至梨形细胞组成。盖生囊状体毛发状近圆柱形,但罕见。所有组织均具锁状联合。
生境:夏秋季散生于草地上,混交林中地上。
世界分布:亚洲(中国);欧洲(丹麦、德国、奥地利、荷兰、英国和法国[模式])。
中国分布:吉林、湖南。
研究标本:吉林:长春市净月潭国家森林公园,2016年8月24日,图力古尔、刘敬,HMJAU44985;长春市吉林农业大学校园,2016年9月8日,图力古尔、刘敬,HMJAU44986;长春市吉林农业大学校园,2016年9月5日,图力古尔、刘敬,HMJAU44987、HMJAU44989;长春市吉林农业大学校园,2016年9月12日,图力古尔、刘敬,HMJAU44992;长春市吉林农业大学校园,2016年7月2日,图力古尔、刘敬,HMJAU44988;安图县二道白河镇,2017年6月30日,图力古尔、刘敬,HMJAU44990;敦化市前进林场,2017年6月26日,图力古尔、刘敬,HMJAU44991;安图县二道白河镇长白山科学院后山,2017年7月25日,图力古尔、刘敬,HMJAU44993;湖南:2022年6月30日,邵阳市南山牧场,宋寒冰、朱力扬,HMJAU62000。
讨论:该种因柄生囊状体为非球颈瓶形被归入具毛组sect. Pilosellae。该种主要特征为菌柄表面被白色粉霜和絮状细小绒毛。该种菌盖颜色以及菌褶着生方式近似于乳白锥盖伞C. apala (Fr.) Arnolds,但前者不自溶,后者自溶,易于区别;该种又与双孢锥盖伞C. bisporigera (Hauskn. & Krisai) Arnolds容易混淆(Arnolds 2005),但后者担孢子大,且呈肾形,易于区分;该种与近缘物种褐纹锥盖伞C. ochrostriata Hauskn. 具相同的孢子大小,但前者干燥后担子体灰色,后者黄褐色,易于区分。
图6
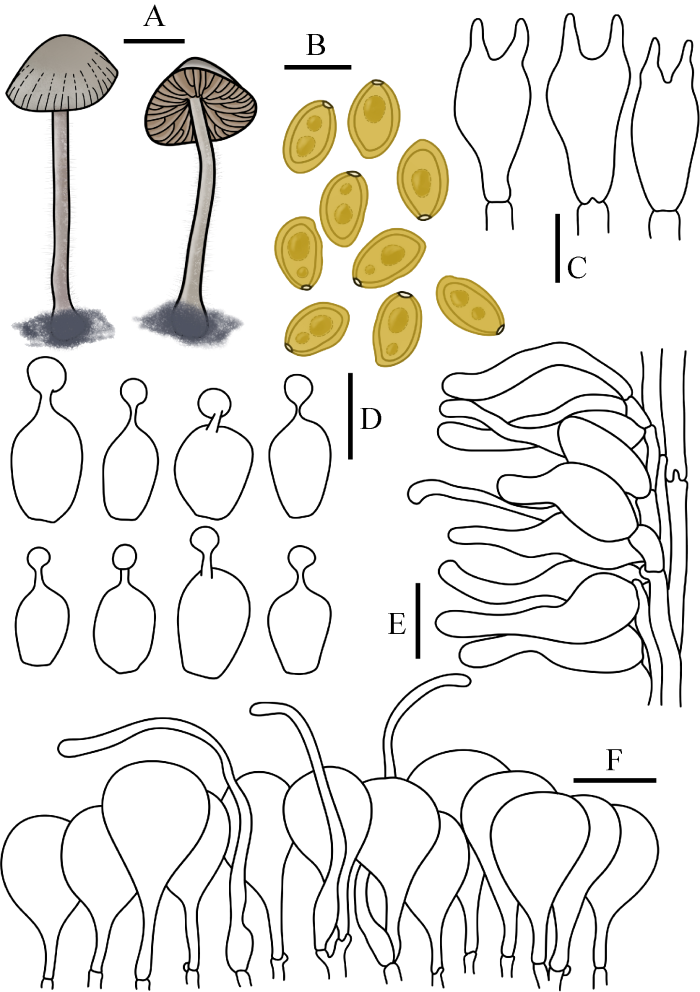
图6
双孢锥盖伞 A:担子体. B:担孢子. C:担子. D:缘生囊状体. E:柄生囊状体. F:菌盖表皮. 标尺:A=1 cm;B-F=10 μm
Fig. 6
Conocybe bisporigera. A: Basidiomata. B: Basidiospores. C: Basidia. D: Cheilocystidia. E: Caulocystidia. F: Pileipellis. Bars: A=1 cm; B-F=10 μm.
Conocybe bisporigera (Hauskn. & Krisai) Arnolds, Persoonia 18(2): 227 (2003)
≡ Conocybe moseri var. bisporigera Hauskn. & Krisa, Persoonia 14(4): 659 (1992)
菌盖直径0.5-2.0 cm,斗笠形,圆锥形,中心灰黑色至深灰黑色,边缘灰色,表面光滑,具明显到中心的沟纹。菌肉薄,无特殊气味。菌褶弯生,稍密,腹鼓状,幅宽1-2 mm,锈褐色。菌柄长3.5-6.5 cm,粗0.5-1.5 mm,圆柱形,基部膨大至球茎状,表面被白色粉霜,具细小绒毛。
担孢子11-13.5×5.5-8.5 μm,Q=1.50-2.27,椭圆形至圆柱形,稍肾形,壁厚,内含油滴,具萌发孔。担子12-24×7-15 μm,棒状,具2担子小梗。缘生囊状体球颈瓶形,14.0-22×7-12 μm,球顶直径3-5 μm。柄生囊状体近圆柱状,烧瓶状,泡囊状。菌盖菌丝膜皮型(“hymeniform”型),由15-29×10-20 μm的球茎细胞和梨形细胞排列而成。盖生囊状体毛发状、烧瓶状、棒状。所有组织均具锁状联合。
生境:秋季散生于草地上。
世界分布:亚洲(中国);欧洲(丹麦、挪威、波兰、德国、奥地利[模式]、荷兰和英国)。
中国分布:吉林。
研究标本:吉林:长春市吉林农业大学校园,2016年9月5日,图力古尔、刘敬,HMJAU45051;长春市吉林农业大学校园,2016年9月6日,图力古尔、刘敬,HMJAU45056;长春市吉林农业大学校园,2016年7月3日,图力古尔、刘敬,HMJAU45057;长春市吉林农业大学校园,2017年7月24日,图力古尔、刘敬,HMJAU45058;安图县二道白河镇池北区白河大街,2017年7月25日,图力古尔、刘敬,HMJAU45055;安图县二道白河镇老山门,2016年8月26日,图力古尔、盖宇鹏、乌日汗、刘敬,HMJAU45059。
讨论:该种因柄生囊状体为非球颈瓶形被归入具毛组sect. Pilosellae。其主要特征为菌盖灰黑色,2担子小梗。该种与条斑锥盖伞C. moseri容易混淆,但前者担子2孢子,担孢子稍肾形,后者担子2或4孢子,从不肾形,易于区分;该种又与近缘物种黑灰锥盖伞C. anthracophila Maire & Kühner ex Kühner & Watling相近,但后者担子4孢子,因此二者易于区分。
锥盖伞属锥盖伞组Conocybe sect. Conocybe分种检索表
1. 菌盖新鲜湿时,以圆锥状,钟状为主······································································2
1’. 菌盖新鲜湿时,以半球状为主············································································10
2. 菌盖圆锥形·····································································································3
2’. 菌盖钟形·····································································································6
3. 菌盖无明显条纹或较浅;担子具4(2)担子小梗················································沙生锥盖伞C. dunensis
3’. 菌盖具条纹;担子具4担子小梗···············································································4
4. 菌褶边缘圆齿状····················································································短柄锥盖伞C. brachypodii
4’. 菌褶边缘光滑·························································································5
5. 具盖生囊状体········································································小孢锥盖伞C. microspora
5’. 无盖生囊状体······························································近柱孢锥盖伞C. cylindricaspora
6. 菌盖淡黄色;孢子稍肾形····································································艾莫锥盖伞C. ammophila
6’. 菌盖褐色至棕色;孢子无肾形··················································································7
7. 菌柄基部具假根··················································································草生锥盖伞C. antipus
7’. 菌柄基部无假根······································································································8
8. 菌褶边缘絮状;柄生囊状体球颈瓶形·····························································里肯锥盖伞C. rickeniana
8’. 菌褶边缘不明显;柄生囊状体具非球颈瓶形元素········································································9
9. 担孢子壁薄;无盖生囊状体········································································大盖锥盖伞C. macrocephala
9’. 担孢子壁厚;有盖生囊状体········································································柔弱锥盖伞C. tenera
10. 菌褶颜色以红棕色为主································································································11
10’. 菌褶颜色以赭石色,褐色为主·························································································13
11. 菌褶边缘略带齿状;菌柄基部球茎不明显················································刺毛锥盖伞C. echinata
11’. 褶缘不明显;菌柄基部明显球茎·························································································12
12. 菌盖中心金黄色至杏黄色;担孢子壁厚········································································黄锥盖伞C. aurea
12’. 菌盖中心深棕色至栗棕色;担孢子壁薄················································近白锥盖伞C. subpallida
13. 菌盖表面具条纹···········································································································14
13’. 菌盖表面无条纹···········································································································16
14. 菌褶具细齿状边缘········································································夏生锥盖伞C. juniana
14’. 菌褶边缘平滑···········································································································15
15. 菌盖表面条纹会消失;孢子稍肾形···················································半球锥盖伞C. semiglobata
15’. 菌盖表面条纹不消失;孢子椭圆形,无肾形 中孢锥盖伞C. mesospora
16. 菌盖无斑纹,担子长度小于25 μm,担孢子明显透镜状············································旱生锥盖伞C. subxerophytica
16’. 菌盖有斑纹,担子长度可达30 μm,担孢子椭圆形···············································拟卵形锥盖伞C. subovalis
图7
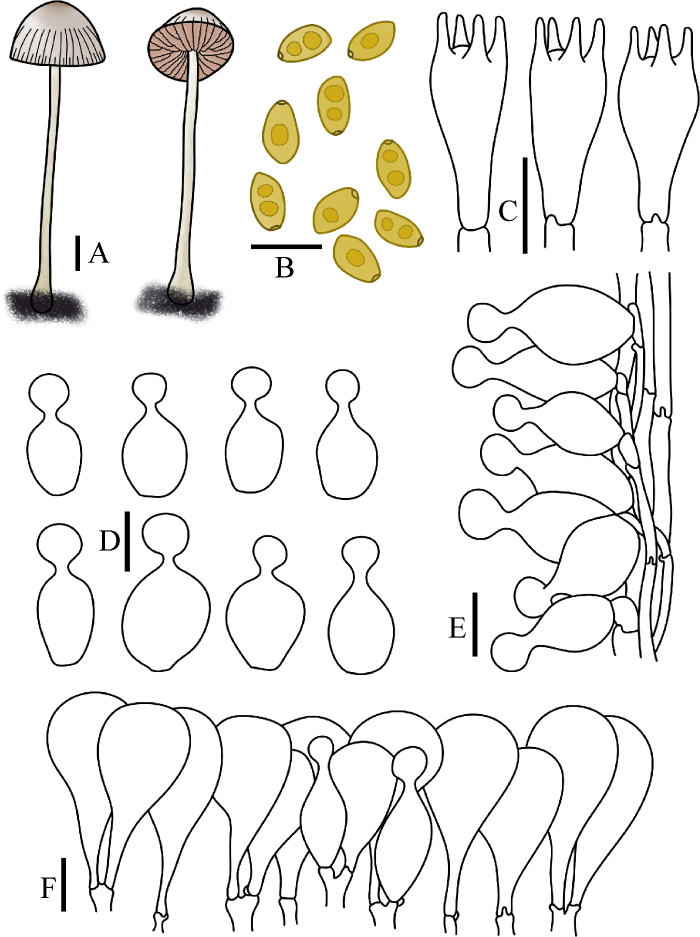
图7
刺毛锥盖伞
A:担子体. B:担孢子. C:担子. D:缘生囊状体. E:柄生囊状体. F:菌盖表皮. 标尺:A=1 cm;B-F=10 μm
Fig. 7
Conocybe echinata.
A: Basidiomata. B: Basidiospores. C: Basidia. D: Cheilocystidia. E: Caulocystidia. F: Pileipellis. Bars: A=1 cm; B-F=10 μm.
Conocybe echinata (Velen.) Singer, Fieldiana, Bot. 21: 103 (1989)
≡ Galera echinata Velen., Novitates Mycologicae Novissimae: 69 (1947)
= Conocybe sordida Kühner & Watling, in Watling, Notes R. bot. Gdn Edinb. 38(2): 339 (1980)
菌盖直径0.8-3.5 cm,初半球形近抛物面形,后凸镜形,中心淡褐色,边缘污白色,表面光滑,水浸状,菌盖边缘具浅沟纹。菌肉黄色至褐色,无特殊气味。菌褶弯生,稍密,幅宽2-4 mm,红褐色,菌褶边缘稍齿状。菌柄长3.0-10.0 cm,粗0.5-2.0 mm,圆柱形,基部球茎状,表面被白色粉霜,具纵向条纹。
担孢子7.5-10×4-6 μm,Q=1.70-1.85,长椭圆形,壁薄,内含油滴,具萌发孔,担孢子在水中呈黄色,KOH溶液中呈淡黄棕色。担子14-25×7-10 μm,棒状,具4担子小梗。缘生囊状体球颈瓶形,17-30×6-9 μm,球顶直径4-6 μm。无侧生囊状体。柄生囊状体球颈瓶形。菌盖菌丝膜皮型(“hymeniform”型),由16-42×10-34 μm的球茎状或梨形细胞栅状排列。盖生囊状体球颈瓶形。所有组织均具锁状联合。
生境:散生或群生在草地、林中和林缘地上。
世界分布:亚洲(中国);欧洲(丹麦、捷克[模式]、挪威、德国、奥地利、荷兰和英国)。
中国分布:吉林。
研究标本:吉林:长春市吉林农业大学校园,2016年5月31日,图力古尔、刘敬,HMJAU45076;长春市吉林农业大学校园,2022年6月26日,图力古尔、王世恩,HMJAU62009。
讨论:Velenovský (1947)选择了种名“echinata”,因为他观察到有刺毛状囊状体,但Singer (1989)和Hausknecht (1999)均未发现此类囊状体,笔者同样也未观察到该类囊状体,若有该类囊状体会在宏观上表现为绒毛存于盖或柄上,但该种囊状体为球颈瓶形,在菌盖上长度小于盖皮细胞,宏观上则呈现菌盖光滑,在菌柄上也仅仅表现为白色粉霜,而中文名则根据拉丁文词源进行命名。其主要特征为菌盖半球形,菌褶红褐色,边缘稍齿状。该种与柔弱锥盖伞C. tenera (Schaeff.) Kühner菌盖形状相似,但后者的担孢子更大,易于区别;该种与近缘物种艾莫锥盖伞C. ammophila M. Lange易混淆,但前者担孢子从不肾形,后者担孢子肾形,可以与之区分。
图8
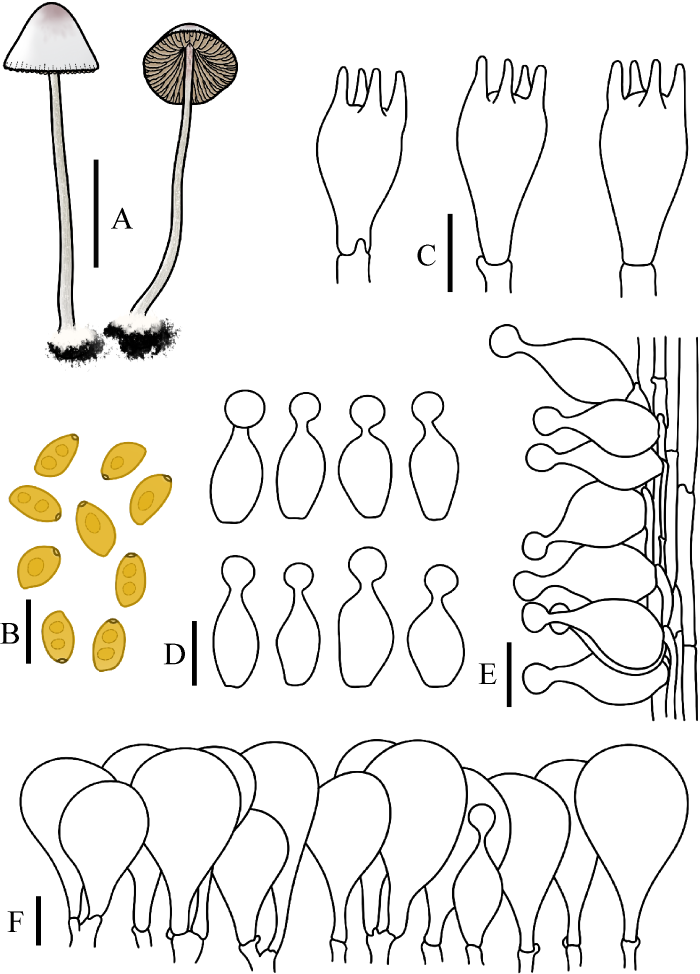
图8
短柄锥盖伞
A:担子体. B:担孢子. C:担子. D:缘生囊状体. E:柄生囊状体. F:菌盖表皮. 标尺:A=1 cm;B-F=10 μm
Fig. 8
Conocybe brachypodii.
A: Basidiomata. B: Basidiospores. C: Basidia. D: Cheilocystidia. E: Caulocystidia. F: Pileipellis. Bars: A=1 cm; B-F=10 μm.
Conocybe brachypodii (Velen.) Hauskn. & Svrček, in Hausknecht, Czech Mycol. 51(1): 43 (1999)
≡ Galera brachypodii Velen., Opera Bot. Ćech. 4: 67 (1947)
= Conocybe macrocephala var. riedheimensis Hauskn. & Enderle, Öst. Z. Pilzk. 9: 95 (2000)
菌盖直径0.5-1.5 cm,高可达1.5 cm,圆锥形近钟形,中心颜色淡棕褐色至红褐色,边缘污白色至浅黄色。菌盖表面光滑,边缘具条纹。菌肉薄,污白色至浅黄色,无特殊气味。菌褶弯生,稍密,不等长,浅黄色至黄褐色,边缘锯齿状。菌柄长1.8-3.8 cm,粗1.0-1.5 mm,圆柱形,基部膨大至球茎状,上端浅棕褐色,下端污白色至黄褐色,表面被白色粉霜,具纵沟纹,球茎状基部具白色绒毛。
担孢子7-9.5×4.5-5.5 μm,Q=1.50-1.80,椭圆形至长椭圆形,壁稍厚,内含油滴,萌发孔中生,孔直径约1 μm,担孢子在水中呈黄色,在KOH溶液中棕褐色。担子15-25×7-10 μm,棒状,具4(2)担子小梗,小梗长1-3 μm,担子具液泡状内含物。缘生囊状体15-28×5-11 μm,球颈瓶形,球顶直径2-6 μm,颈长1-6 μm。侧生囊状体无。柄生囊状体16-32×7-14 μm,球颈瓶形,球顶直径3-6 μm,颈长2-8 μm。菌盖菌丝膜皮型(“hymeniform”型),由16-43×10-22 μm的球茎或倒梨形细胞排列而成。盖生囊状体球颈瓶形。所有组织均具锁状联合。
生境:散生于草地、林地上。
世界分布:亚洲(中国);欧洲(丹麦、挪威、俄罗斯、捷克[模式]、德国、奥地利、荷兰、英国和法国)。
中国分布:黑龙江、吉林。
研究标本:吉林:吉林农业大学校园,2016年9月5日,图力古尔、刘敬,HMJAU45017;吉林农业大学校园,2022年8月30日,图力古尔、宋寒冰,HMJAU64927,HMJAU64929;吉林农业大学校园,2022年8月31日,图力古尔、宋寒冰,HMJAU64928。
讨论:该种因其柄囊体为球颈瓶形被归入锥盖伞组sect. Conocybe。该种与柔弱锥盖伞C. tenera菌盖大小及颜色相近,但前者担孢子长度小于10 μm,后者担孢子长度大于10 μm,易于区别;该种又与中孢锥盖伞C. mesospora Kühner ex Watling易混淆,但前者菌盖棕褐色至污白色,菌褶边缘锯齿状,与氨水呈弱阳性反应,后者菌盖颜色橙色到棕橙色,菌褶边缘平滑,与氨水呈阴性反应,因此二者易于区分。
锥盖伞属混杂组Conocybe sect. Mixtae分种检索表
1. 菌盖新鲜湿时,钟状,顶针状,菌盖高大于宽或等同于宽·······························································2
1’. 菌盖新鲜湿时,圆锥形,近半球形或中部凸起,菌盖宽大于高········································4
2. 无盖生囊状体················································································复囊体锥盖伞C. digitalina
2’. 有盖生囊状体···············································································································3
3. 担孢子长度小于20 μm································································································绒柄锥盖伞C. pubescens
3’. 担孢子长度可达24 μm··························································································大孢锥盖伞C. macrospora
4. 未观察到锁状联合;无盖生囊状体·····················································································小脆锥盖伞C. fragilis
4’. 具锁状联合;有盖生囊状体··································································································5
5. 菌褶具絮状边缘;担子2孢子················································································褐柄锥盖伞C. ambigua
5’. 菌褶边缘平滑;担子4孢子·············································································近绒柄锥盖伞C. subpubescens
小鳞伞属Pholiotina分组检索表
1. 担孢子表面粗糙或皱纹、微小的疣·························································疣孢组sect. Verrucisporae
1’. 担孢子表面光滑·········································································2
2. 菌幕残留·················································································3
2’. 菌幕不残留··············································································5
3. 缘生囊状体球颈瓶形·······································································过渡组sect. Intermediae
3’. 缘生囊状体非球颈瓶形········································································4
4. 菌幕残留在菌柄上呈膜质环状·································································小鳞伞组sect. Pholiotina
4’. 菌幕残留在菌盖上呈絮状或残片,从不成环········································································被外菌幕组sect. Vestitae
5. 菌盖或菌柄基部伤后呈蓝色或蓝绿色调···········································································蓝色组sect. Cyanopodae
5’. 菌盖或菌柄基部无蓝色或蓝绿色调·············································································被毛组sect. Piliferae
小鳞伞属疣孢组Pholiotina sect. Verrucisporae分种检索表
1. 担子4孢子;缘生囊状体大部分烧瓶形·················································糙孢小鳞伞Ph. dasypus
1’. 担子4和2孢子混合;缘生囊状体大部分椭圆形 ···········································泡囊小鳞伞Ph. utriformis
图9
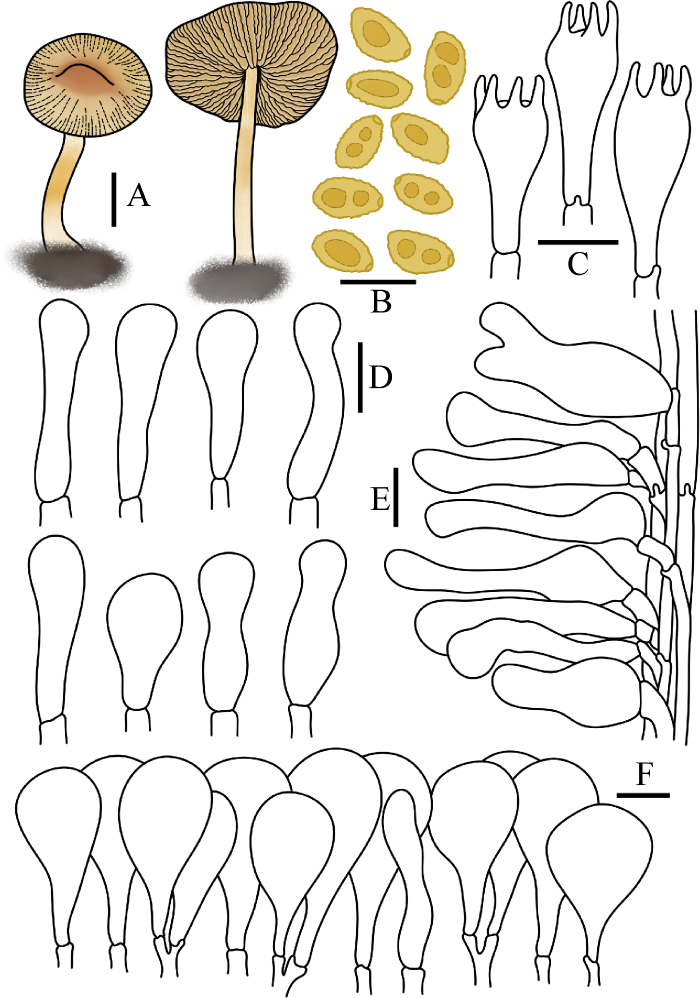
图9
糙孢小鳞伞
A:担子体. B:担孢子. C:担子. D:缘生囊状体. E:柄生囊状体. F:菌盖表皮. 标尺:A=1 cm;B-F=10 μm
Fig. 9
Pholiotina dasypus.
A: Basidiomata. B: Basidiospores. C: Basidia. D: Cheilocystidia. E: Caulocystidia. F: Pileipellis. Bars: A=1 cm; B-F=10 μm.
Pholiotina dasypus (Romagn.) P.-A. Moreau, Fungal Diversity 20: 135 (2005)
≡ Naucoria dasypus Romagn., Bull. trimest. Soc. mycol. Fr. 53: 121 (1937)
≡ Hebeloma dasypus (Romagn.) Singer, Lilloa 22: 538 (1951) [1949]
菌盖直径1.5-3.5 cm,初期钟形近圆锥形,后期中部凸起,中心红褐色,边缘淡褐色,菌盖表面稍粘,水浸状,光滑或稍褶皱,具明显沟纹。菌肉薄,淡褐色,无特殊气味。菌褶弯生,密,腹鼓状,褶幅2-4 mm,菌褶边缘具白色小齿。菌柄长4.0-7.0 cm,直径1.0-4.0 mm,圆柱形,基部明显膨大,上端浅黄色至黄褐色,下端污白色,表面被白色粉霜,稍具纵沟纹。
担孢子7.5-11×4-6 μm,Q=1.6-2.5,椭圆形至圆柱形,光学显微镜下略粗糙,扫描电镜下表面具小疣凸,壁薄,内含油滴,具萌发孔。担子9-24×7-10 μm,棒状,具4担子小梗,基部具锁状联合。缘生囊状体21-34×4-13 μm,大部分烧瓶形,部分泡囊形或棒状。无侧生囊状体。柄生囊状体24-48×7-10 μm,近烧瓶形。菌盖菌丝膜皮型(“hymeniform”型),由29-47×7-17 μm的球茎细胞或短梨形细胞排列而成,基部具锁状联合。盖生囊状体与缘生囊状体相似,但罕见。
生境:秋季单生于林下枯枝落叶层中碎木片上、草地上。
世界分布:亚洲(中国);欧洲(丹麦、波兰、德国、奥地利、比利时和法国[模式]);北美洲(美国)。
中国分布:吉林。
研究标本:吉林:长春市净月潭国家森林公园,2017年8月10日,图力古尔、刘敬,HMJAU45095、HMJAU45096;长春市净月潭国家森林公园,2017年7月7日,图力古尔、刘敬,HMJAU 45099、HMJAU 45102、HMJAU 45103;长春市净月潭国家森林公园,2017年7月18日,图力古尔、刘敬,HMJAU 45100、HMJAU 45104、HMJAU 45105;安图县二道白河镇天伦公园,2016年6月16日,图力古尔、刘敬,HMJAU45098;长春市吉林农业大学校园,2022年5月30日,图力古尔,HMJAU62002;长春市吉林农业大学校园,2022年5月31日,图力古尔、宋寒冰,HMJAU62003。
讨论:该种因担孢子粗糙具疣凸被归入疣孢组sect. Verrucisporae。其主要特征为担孢子粗糙,具疣凸。该种与南方小鳞伞Ph. australis (Singer) Watling具相同的粗糙孢子,我国尚未发现后者的分布,易于区分;该种又与泡囊小鳞伞Ph. utriformis (P.D. Orton) Bon易混淆,Hausknecht (2009)认为该种与后者同物异名,但后者担子4和2孢子混合,缘生囊状体椭圆形,可与该种区分;该种与其缘物种沟纹小鳞伞Ph. sulcata Arnolds & Hauskn系统发育上相近,但后者孢子光滑,因此二者易于区分。
小鳞伞属过渡组Pholiotina sect. Intermediae分种检索表
1. 菌盖半球形,橙色至黄褐色;具盖生囊状体··············································纤丝小鳞伞Ph. fibrillosipes
1’. 菌盖凸镜形,钝圆锥形,暗红褐色,巧克力色;无盖生囊状体··············································褐小鳞伞Ph. brunnea
小鳞伞属小鳞伞组Pholiotinasect.Pholiotina分种检索表
1. 菌盖红棕色调为主····································································································2
1’. 菌盖以褐色调和赭色调为主····························································································4
2. 褶缘具小齿············································································前小鳞伞Ph. percincta
2’. 褶缘平滑···············································································································3
3. 担孢子壁薄;锁状联合常见·························································皱盖小鳞伞Ph. rugosa
3’. 担孢子壁厚;锁状联合罕见·························································近柱囊小鳞伞Ph. utricystidiata
4. 生于苔藓层和针叶林·························································线小鳞伞Ph. filaris
4’. 生于落叶林和混交林·········································································································5
5. 菌褶边缘稍齿状;担孢子壁薄······················································阿伦尼小鳞伞Ph. arrhenii
5’. 菌褶具絮状物;担孢子壁厚······················································脆盖小鳞伞Ph. vexans
小鳞伞属被外菌幕组Pholiotina sect. Vestitae分种检索表
1. 菌盖凸镜形;菌褶红褐色····················································林间小鳞伞Ph. nemoralis
1’. 菌盖圆锥形到中部凸起;菌褶橙棕色····················································2
2. 菌盖上菌幕残余锯齿状;担孢子无萌发孔····················································齿缘小鳞伞Ph. serrata
2’. 菌盖上菌幕残余絮状;担孢子萌发孔小····················································多变小鳞伞Ph. mutabilis
齿缘小鳞伞
Pholiotina serrata (T. Bau & J. Liu) T. Bau et H. B. Song, comb. nov.
MycoBank: MB 845742
≡ Conocybe serrata T. Bau & J. Liu, in Liu & Bau, Phytotaxa 357(4): 267 (2018)
齿缘小鳞伞Ph. serrata缘生囊状体非球颈瓶形,属于小鳞伞属Pholiotina物种;菌幕残留在菌盖边缘,不在菌柄上呈环,应被划入被外菌幕组sect. Vestitae,且形态学分组和系统发育结果一致,证明其为小鳞伞。
The cheilocystidia of Ph. serrata is non-lecythiform. The veil remains on the edge of the pileus, and is not in a ring on the stipe, so the species should be classified in sect. Vestita. Consistency of morphological grouping and phylogenetic results proved that this species belonged to Pholiotina.
小鳞伞属蓝色组Pholiotina sect. Cyanopodae分种检索表
1. 褶缘不明显;担孢子萌发孔略偏心····················································蓝柄小鳞伞Ph. cyanopus
1’. 褶缘絮状或细小圆齿;担孢子萌发孔小····················································史密斯小鳞伞Ph. smithii
图10
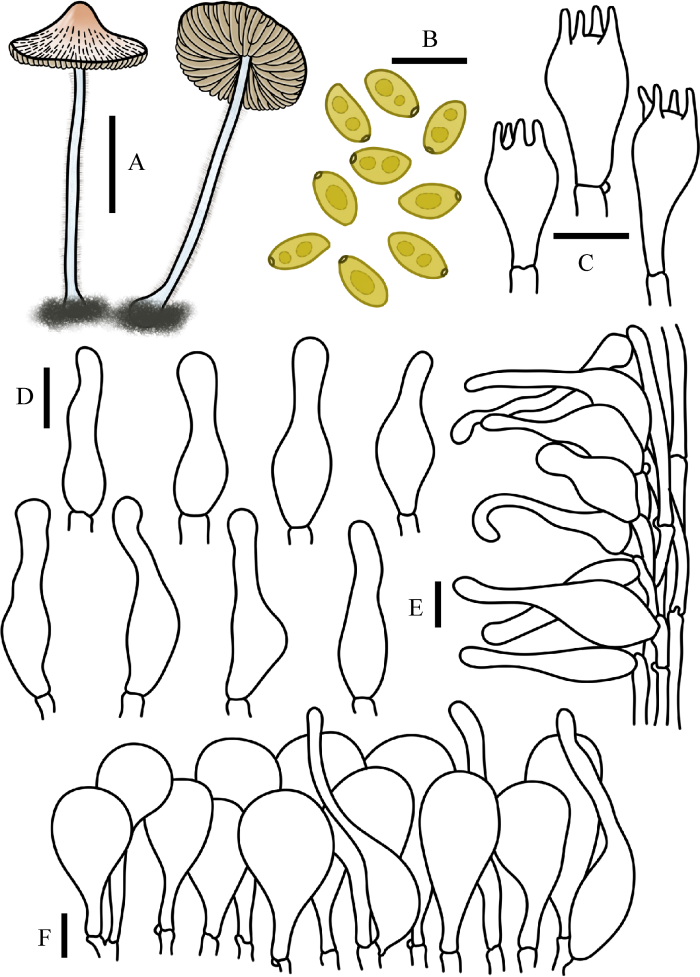
图10
蓝柄小鳞伞
A:担子体. B:担孢子. C:担子. D:缘生囊状体. E:柄生囊状体. F:菌盖表皮. 标尺:A=1 cm;B-F=10 μm
Fig. 10
Pholiotina cyanopus.
A: Basidiomata. B: Basidiospores. C: Basidia. D: Cheilocystidia. E: Caulocystidia. F: Pileipellis. Bars: A=1 cm; B-F=10 μm.
Pholiotina cyanopus (G.F. Atk.) Singer, Trudy Bot. Inst. Akad. Nauk SSSR, ser. 2, Sporov. Rast. 6: 425 (1950)
≡ Galerula cyanopus G.F. Atk., Proc. Am. phil. Soc. 57: 367 (1918)
≡ Conocybe cyanopus (G.F. Atk.) Kühner [as 'cyanopoda'], Encyclop. Mycol. 7: 128 (1935)
菌盖直径0.5-1.5 cm,初期圆锥形,后渐中部凸起,中心赭橙色,边缘淡黄褐色,菌盖表面水浸状,具明显沟纹。菌肉薄,淡黄色,无特殊气味。菌褶弯生,稍密,腹鼓状,黄褐色,褶幅1-2 mm。菌柄长3.0-6.0 cm,粗0.5-1.0 mm,圆柱形,纤维质,基部膨大至球茎状,表面被白色粉霜,细小绒毛,污白色,伤变蓝。
担孢子7.5-9.5×4-5 μm,Q=1.60-1.90,椭圆形至长椭圆形,薄壁,光滑,内含油滴,萌发孔中生至稍偏心。担子14-24×7-10 μm,棒状,具4担子小梗,基部具锁状联合。缘生囊状体19-35×4-10 μm,近烧瓶形,泡囊形,棒状。无侧生囊状体。柄生囊状体与缘生囊状体相似。菌盖菌丝膜皮型(“hymeniform”型),由39-54× 14-22 μm的球茎细胞和短梨形细胞栅状排列而成,具锁状联合。盖生囊状体毛发状至烧瓶形。
生境:秋季单生于林下枯枝落叶层中碎木片上和路旁草地上。
世界分布:亚洲(中国、俄罗斯);欧洲(丹麦、挪威、瑞典、德国、瑞士、荷兰和法国);北美洲(美国[模式])。
中国分布:吉林。
研究标本:吉林:长春市净月潭国家森林公园,2017年8月10日,图力古尔、刘敬,HMJAU45092、HMJAU45096;安图县二道白河镇天伦公园,2016年6月16日,图力古尔、刘敬,HMJAU45098;长春市净月潭国家森林公园鹿苑附近,2021年9月2日,图力古尔、朱力扬、宋寒冰,HMJAU62007。
讨论:该种因其菌柄伤变蓝而被归入蓝色组sect. Cyanopodae。该种偶尔与糙孢小鳞伞Ph. dasypus生在一处,易混淆(Arnolds 2005),但后者伤不变色,易于区分;该种与近缘物种史密斯小鳞伞Ph. smithii (Watling) Enderle在分子系统上相近,均伤变蓝,但后者菌盖钝圆锥至抛物面形,菌褶具细小圆齿,因此二者可以区分。
小鳞伞属被毛组Pholiotina sect. Piliferae分种检索表
1. 菌盖半球形至凸镜形,以白色调为主··································································喜粪小鳞伞Ph. coprophila
1’. 菌盖圆锥状,以红色调为主··········································································2
2. 菌褶略带细齿状边缘;无盖生囊状体·····································································沟纹小鳞伞Ph. sulcata
2’. 菌褶边缘絮状;有盖生囊状体··········································································易碎小鳞伞Ph. aberrans
图11
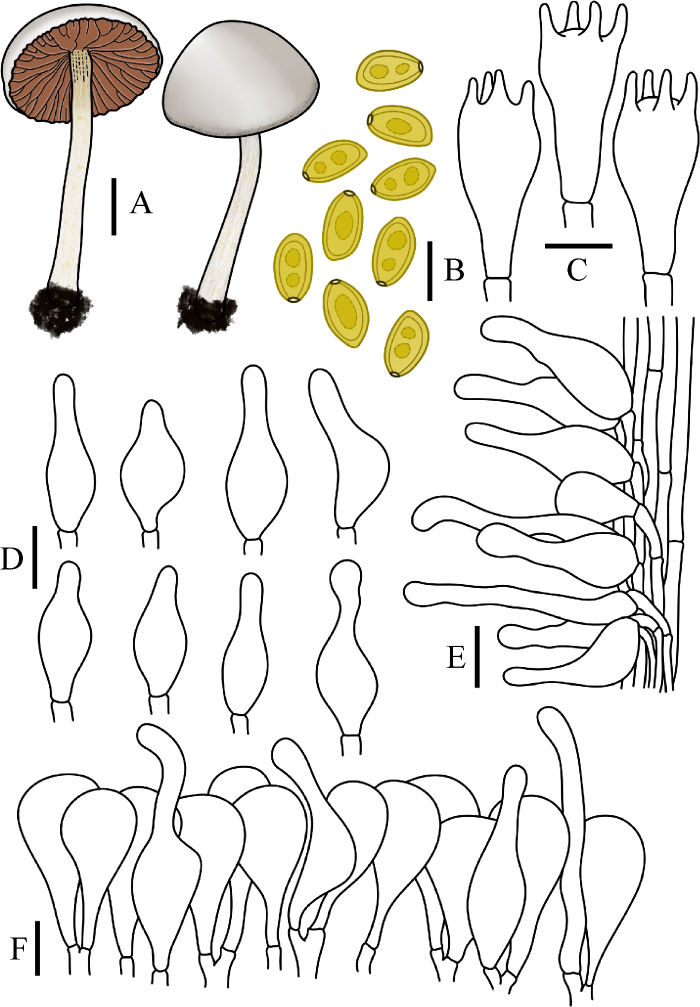
图11
喜粪小鳞伞
A:担子体. B:担孢子. C:担子. D:缘生囊状体. E:柄生囊状体. F:菌盖表皮. 标尺:A=1 cm;B-F=10 μm
Fig. 11
Pholiotina coprophila.
A: Basidiomata. B: Basidiospores. C: Basidia. D: Cheilocystidia. E: Caulocystidia. F: Pileipellis. Bars: A=1 cm; B-F=10 μm.
Pholiotina coprophila (Kühner) Singer, Acta Inst. bot. Komarov. Acad. Sci., Pl. Crypt, ser. 2 6: 434 (1950)
≡ Galera coprophila Kühner, Botaniste 17(1-4): 169 (1926)
≡ Conocybe coprophila (Kühner) Kühner, Encyclop. Mycol. 7: 125 (1935)
菌盖直径0.8-3.2 cm,半球形至凸镜形,奶油白色至浅黄色,菌盖表面稍粘,无沟纹或沟纹不明显。菌肉薄,灰白色,无特殊气味。菌褶直生近弯生,腹鼓状,幅宽1-3 mm,红褐色,菌褶边缘絮状。菌柄长4.0-7.0 cm,粗2.0-3.0 mm,圆柱形,基部稍膨大,污白色至浅赭色,表面被白色粉霜,具纵条纹。
担孢子7.5-13.5×5-6.5 μm,Q=1.50-2.30,椭圆形至圆柱形,壁厚,内含油滴,具明显萌发孔。担子21-30×9-15 μm,棒状,具4担子小梗,未见锁状联合。缘生囊状体21-32×6-13 μm,近烧瓶形,纺锤形至披针形。无侧生囊状体。柄囊体形状与缘生囊状体相近,19-42×4-15 μm,近烧瓶形,纺锤形至披针形。菌盖菌丝膜皮型(“hymeniform”型),由21-37×12-17 μm的球茎细胞和梨形细胞排列而成,锁状联合未见。盖生囊状体烧瓶形,近圆柱形。
生境:秋季单生或散生于云杉林草地中粪上、沙地樟子松林中粪上。
世界分布:亚洲(中国);欧洲(冰岛、丹麦、挪威、瑞典、奥地利、瑞士、荷兰和法国[模式]);大洋洲(澳大利亚)。
中国分布:吉林、内蒙古和新疆。
研究标本:吉林:长春市吉林农业大学校园,2016年9月8日,图力古尔、刘敬,HMJAU45087;内蒙古:鄂温克旗辉河保护区,2016年9月5日,盖宇鹏、颜俊清、刘晓亮、石书锋,HMJAU45088、HMJAU48089;新疆:喀纳斯风景区,2021年8月15日,叶芊歧,HMJAU62008。
讨论:该种因缘生囊状体烧瓶形近披针形而被划入被毛组sect. Piliferae。其主要特征为菌盖表面稍粘,粪生,无菌幕残留,菌柄伤不变色。该种在系统学位置上与蓝色组sect. Cyanopodae分支互为姊妹类群,但后者中物种伤变蓝,因此二者易于区分。
3 讨论
根据Tóth et al. (2013)的系统发育框架,结合Hausknecht (2009)的形态分类观点,加入我国材料的基因序列,重建了基于ITS1、5.8S和ITS2的系统发育树,显示锥盖伞属Conocybe为单系,小鳞伞属Pholiotina为多系。基于我国文献记载和标本的研究,采纳Kühner (1935)、Arnolds (2005)和Hausknecht (2009)的观点,明确我国锥盖伞属Conocybe和小鳞伞属Pholiotina划分界线。本研究认为锥盖伞属Conocybe是粪锈伞科Bolbitiaceae中的高级进化类群,缘生囊状体均为球颈瓶形至保龄球形,无菌幕残留,菌柄纤柔,子实下层菌褶菌髓被压缩成窄圆柱形或近圆柱形,子实层基部菌髓气泡状;小鳞伞属Pholiotina是结合了3个分支的特征,其菌褶菌髓气泡状近宽柱状,菌髓中心不被压缩,缘生囊状体非球颈瓶形(泡囊状、近圆柱状、披针形和烧瓶形),若为球颈瓶形则带菌幕残留,如过渡组sect. Intermediae中的褐小鳞伞Ph. brunnea (J.E. Lange & Kühner ex Watling) Singer和中间小鳞伞Ph. intermedia (Kühner) Singer及其他物种,其均具球颈瓶形囊状体,但褐小鳞伞Ph. brunnea菌盖具菌幕残留,中间小鳞伞Ph. intermedia菌幕在柄上呈环,其均属于被外菌幕组sect. Vestitae分支的末端,暗示了缘生囊状体是从非球颈瓶形向球颈瓶形方向演化的;再如,小鳞伞组sect. Pholiotina中的末端为近柱囊小鳞伞Ph. utricystidiata,其缘生囊状体近球颈瓶形,但不完全是球颈瓶形,处于一种过渡态,这是一种趋同演化的现象。同理,Pholiotina Glade 1与锥盖伞属Conocybe互为姊妹类群,亲缘关系近,其中喜粪小鳞伞Ph. coprophila和蓝柄小鳞伞Ph. cyanopus等物种缘生囊状体为烧瓶形,近披针形,有时长颈顶端会稍膨大呈球状,这是球颈瓶形的前体,处于中间态,进一步证明了缘生囊状体是从非球颈瓶形向球颈瓶形方向演化的,从而也说明了缘生囊状体是划分锥盖伞属Conocybe和小鳞伞属Pholiotina的重要特征,本研究认为这同样是造成这些年锥盖伞属和小鳞伞属界限不明确的原因,在小鳞伞组sect. Pholiotina中物种进化末端会演化成球颈瓶形囊状体,但却忽略了这些物种具菌幕残留,属于小鳞伞属Pholiotina物种。本研究认为造成小鳞伞属Pholiotina多系的原因是Pholiotina Glade 1和Pholiotina Glade 2均处于演化的中间过渡态,缘生囊状体形态相近,其次是这2个分支物种的材料较少,以及前期分子生物学手段可能还不够完善。
对我国锥盖伞属和小鳞伞属下划分,采纳的是Hausknecht (2009)的形态学分类观点,将我国锥盖伞属Conocybe划分为5个属下等级,白色组sect. Candidae,担子体成熟后菌盖菌褶多自溶,存在拟担子(周细胞),类似于鬼伞,有时难以区分,容易混淆,属于弥散性现象(朱力扬等 2022),在该组中乳白锥盖伞C. apala的种加词有纤弱、柔弱的意思,Hausknecht (2009)认为乳白锥盖伞C. apala与白柄锥盖伞C. albipes不同,可通过拟担子是否膨大进行区分,1998年将皱锥盖伞C.crispa作为白柄锥盖伞C. albipes的一个变种处理(Hausknecht 1998),但笔者并未采集过白柄锥盖伞C. albipes和皱锥盖伞C.crispa,也未借阅过标本,且皱锥盖伞C. crispa的第一份标本已经丢失,仍然存疑(Hausknecht & Krisai-Greilhuber 2004),对于乳白锥盖伞C. apala与白柄锥盖伞C. albipes是否为同物异名的问题,笔者发现乳白锥盖伞C. apala存在自溶与不自溶的现象,且基于ITS的分子结果完全相同,宏观形态和显微特征也几乎一致,不自溶的常生于牛粪上,具自溶特征的常生于草地,由此笔者有两种猜测,其一,乳白锥盖伞C. apala与白柄锥盖伞C. albipes为同物异名,因生境不同所导致的是否自溶,其二,乳白锥盖伞C. apala与白柄锥盖伞C. albipes为不同种,但存在复合群的问题,暂时难以区分,需后续深入研究。而乳白锥盖伞C. apala与柔弱锥盖伞C. tenera为不同种,不仅通过ITS可以区分,其宏观特征也可被归入不同组中,因其余种与C. apala的关系存疑,所以用乳白锥盖伞防止混淆;赭小皮伞组sect. Ochromarasmius,孢子粗糙,具疣状点或褶皱,中国仅记载一个物种为灌丛锥盖伞C. dumetorum (Velen.) Svrček (臧穆 1996),其在系统发育结果中的近缘物种草生锥盖伞C. antipus (Lasch) Kühner和卡氏锥盖伞C. karinae Gubitz & Hauskn.均不属于该组物种,形态和系统不能良好拟合,需进一步研究;具毛组sect. Pilosellae,柄囊体非球颈瓶形,属于锥盖伞属Conocybe系统发育的基部,其形态划分与系统发育结果一致;锥盖伞组sect. Conocybe和混杂组sect. Mixtae之间的划分存在很多问题,物种界限不是很清晰,序列拓扑结构之间存在嵌套现象,仅依靠柄囊体的形态不足以完美划分,还需寻找新的足够区分的物种特征,图1中锥盖伞组sect. Conocybe和混杂组sect. Mixtae中蓝色字体的物种均存在嵌套现象;且我国尚未发现小辛格组sect. Singerella中的物种。
将我国小鳞伞属划分为6个属下等级,传统的小鳞伞属Pholiotina是Pholiotina Glade 3,具菌幕残留,其中包含3个组,小鳞伞组sect. Pholiotina,菌幕在菌柄上成环,线小鳞伞Ph. filaris (Fr.) Singer具菌环,但拓扑结构没有与小鳞伞组sect. Pholiotina汇聚在一支,汇聚在另一支的支持率也很低,笔者在手动调矩阵时发现其序列简并碱基较多,长度和质量存在欠缺。对于我国已记载物种近柱囊锥盖伞C. utricystidiata、阿伦尼锥盖伞C. arrhenii (Fr.) Kits van Wav.、前锥盖伞C. percincta P.D. Orton、皱盖锥盖伞C. rugosa (Peck) Watling和线锥盖伞C. filaris均具菌环,缘生囊状体为非球颈瓶形,且具小鳞伞属Pholiotina的同义词,已作为小鳞伞属物种处理(Li & Azbukina 2011;王薇 2014;刘敬 2018);被外菌幕组sect. Vestitae,菌幕残留在菌盖边缘呈絮状或齿状,齿缘小鳞伞Ph. serrata菌盖边缘具齿状菌幕,无菌环,系统发育结果也在被外菌幕组sect. Vestitae中,所以将其作为新组合(Liu & Bau 2018),而我国记载的林间锥盖伞C. nemoralis和多变锥盖伞C. mutabilis同样具该组的特征和具小鳞伞属Pholiotina的同义词,故将其作为小鳞伞属物种处理(Li & Azbukina 2011;冶晓燕 2021);过渡组sect. Intermediae,缘生囊状体球颈瓶形,具菌幕残留,菌幕残留于菌盖或于菌柄成环,我国记载的褐锥盖伞C. brunnea和纤丝锥盖伞C. fibrillosipes已被划入该组,但纤丝小鳞伞Ph. fibrillosipes未能收集到标本材料,曾经也没有小鳞伞属Pholiotina的同义词,但其具菌幕残余,应为小鳞伞属物种,甚至Horak & Hausknecht (2002)认为纤丝锥盖伞C. fibrillosipes 是来自欧洲的褐小鳞伞Ph. brunnea 的同义词,所以本研究未将其进行组合,只是作为小鳞伞属物种处理(Li & Azbukina 2011)。在Pholiotina Glade 2中,疣孢组sect. Verrucisporae担孢子表面粗糙或皱纹,有微小的疣,主要包括糙孢小鳞伞Ph. dasypus和泡囊小鳞伞Ph. utriformis,Hausknecht (2009)认为其为同物异名,本文也已讨论,见糙孢小鳞伞Ph. dasypus描述下的讨论。Pholiotina Glade 1中蓝色组sect. Cyanopodae,菌柄伤后具蓝绿色调,主要包括蓝柄小鳞伞Ph. cyanopus和史密斯小鳞伞Ph. smithii;而沟纹小鳞伞Ph. sulcata、喜粪小鳞伞Ph. coprophila和易碎小鳞伞Ph. aberrans (Kühner) Singer因孢子光滑,菌柄菌盖无蓝绿色调,缘生囊状体非球颈瓶形,不规则烧瓶形近披针状而被划入被毛组sect. Piliferae,但其物种没有汇聚在一支,还需重新确定分类特征,将其划分。
我国记载的大部分种是作为锥盖伞属Conocybe物种来处理,本研究虽然已对我国锥盖伞属Conocybe和小鳞伞属Pholiotina进行划分,但还有很多问题需要解决,如小鳞伞属Pholiotina多系来源的问题,以及该两属一些物种形态学划分与系统发育结果不能拟合。尽管我国对大型森林真菌的多样性进行了系统性研究,并且发表了诸多新分类单元(戴玉成等2021),但这些前期研究几乎没有涉及森林生态系统中一些个体较小的蘑菇。本研究报道了我国锥盖伞属37种、小鳞伞属18种,是对我国森林大型真菌的补充,也说明今后类似类群的多样性还需要进一步研究。
参考文献
Bolbitiaceae
List of agarics and boletoid fungi from eastern Inner Mongolia (Ⅱ)
A revised checklist of poisonous mushrooms in China
Notes on Basidiomycetes of Jilin Province (Ⅲ)
Agarics and other macrobasidiomycetes from Ussuri River Valley
Macrofungus flora of Guangdong Province
Diversity and systematics of the important macrofungi in Chinese forests
Conocybe-Pholiotina-Studien I: bestimmungsschlüssel für die europäischen Arten der Gattung Conocybe Fayod
Studies in Conocybe and Pholiotina VI: description of finds
Studies in Conocvbe and Pholiotina VIII
Prodrome d'une histoire naturelle des Agaricinés
Systema mycologicum, sistens fungorum ordines, genera et species
Epicrisis systematis mycologici seu synopsis hymenomycetum
Psilocybin produces substantial and sustained decreases in depression and anxiety in patients with life-threatening cancer: a randomized double-blind trial
Cancer patients often develop chronic, clinically significant symptoms of depression and anxiety. Previous studies suggest that psilocybin may decrease depression and anxiety in cancer patients. The effects of psilocybin were studied in 51 cancer patients with life-threatening diagnoses and symptoms of depression and/or anxiety. This randomized, double-blind, cross-over trial investigated the effects of a very low (placebo-like) dose (1 or 3 mg/70 kg) vs. a high dose (22 or 30 mg/70 kg) of psilocybin administered in counterbalanced sequence with 5 weeks between sessions and a 6-month follow-up. Instructions to participants and staff minimized expectancy effects. Participants, staff, and community observers rated participant moods, attitudes, and behaviors throughout the study. High-dose psilocybin produced large decreases in clinician- and self-rated measures of depressed mood and anxiety, along with increases in quality of life, life meaning, and optimism, and decreases in death anxiety. At 6-month follow-up, these changes were sustained, with about 80% of participants continuing to show clinically significant decreases in depressed mood and anxiety. Participants attributed improvements in attitudes about life/self, mood, relationships, and spirituality to the high-dose experience, with >80% endorsing moderately or greater increased well-being/life satisfaction. Community observer ratings showed corresponding changes. Mystical-type psilocybin experience on session day mediated the effect of psilocybin dose on therapeutic outcomes.ClinicalTrials.gov identifier: NCT00465595.© The Author(s) 2016.
New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0
DOI:10.1093/sysbio/syq010
PMID:20525638
[本文引用: 1]

PhyML is a phylogeny software based on the maximum-likelihood principle. Early PhyML versions used a fast algorithm performing nearest neighbor interchanges to improve a reasonable starting tree topology. Since the original publication (Guindon S., Gascuel O. 2003. A simple, fast and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 52:696-704), PhyML has been widely used (>2500 citations in ISI Web of Science) because of its simplicity and a fair compromise between accuracy and speed. In the meantime, research around PhyML has continued, and this article describes the new algorithms and methods implemented in the program. First, we introduce a new algorithm to search the tree space with user-defined intensity using subtree pruning and regrafting topological moves. The parsimony criterion is used here to filter out the least promising topology modifications with respect to the likelihood function. The analysis of a large collection of real nucleotide and amino acid data sets of various sizes demonstrates the good performance of this method. Second, we describe a new test to assess the support of the data for internal branches of a phylogeny. This approach extends the recently proposed approximate likelihood-ratio test and relies on a nonparametric, Shimodaira-Hasegawa-like procedure. A detailed analysis of real alignments sheds light on the links between this new approach and the more classical nonparametric bootstrap method. Overall, our tests show that the last version (3.0) of PhyML is fast, accurate, stable, and ready to use. A Web server and binary files are available from http://www.atgc-montpellier.fr/phyml/.
Beiträge zur Kenntnis der Bolbitiaceae 1. Pholiotina subnuda und Conocybe hexagonospora
Beiträge zur Kenntnis der Bolbitiaceae 2. Die Variabilität von Conocybe dumetorum
Beiträge zur Kenntnis der Bolbitiaceae 3. Europäische Conocybe-Arten mit wurzelndem oder tief im Substrat eingesenktem Stiel
Beitrage zur Kenntnis der Bolbitiaceae 4. Die Sektion Candidae und andere hellhutige Arten der Gattung Conocybe
Beiträge zur Kenntnis der Bolbitiaceae 5. Die Conocybe rickeniana-und C. magnicapitata- Gruppe in Europa
Beitrage zur Kenntnis der Bolbitiaceae 7. Die Conocybe tenera-Gruppe, Teil 2, und eine Revision der Arten um Conocybe mesospora in Europa
Conocybe tuxlaensis und C
Beiträge zur Kenntnis der Bolbitiaceae 9. Conocybe Sekt
Beiträge zur Kenntnis der Bolbitiaceae 10. Conocybe Sektion Pilosellae
Monograph of the genera Conocybe Fayod, Pholiotina Fayod in Europe
Interesting species of Conocybe (Agaricales, Bolbitiaceae) from Gallura (NE Sardinia, Italy)
Conocybe-Pholiotina- Studien III. Drei neue Conocybe-Arten aus Italien
The genera Conocybe and Pholiotina (Agaricomycotina, Bolbitiaceae) in temperate Asia
Schwarzhütige Conocybe-Arten
Type studies in north American species of Bolbitiaceae belonging to the genera Conocybe and Pholiotina
Infrageneric division of the genus Conocybe-a classical approach
Infrageneric division of the genus Pholiotina-a classical approach
Two new Conocybe species from Europe and corrections to the monograph Conocybe-Pholiotina
Two remarkable species of Bolbitiaceae (Agaricales) from Croatia
Phylogenetic relationships in the mushroom genus Coprinus and dark-spored allies based on sequence data from the nuclear gene coding for the large ribosomal subunit RNA: divergent domains, outgroups, and monophyly
DOI:10.1006/mpev.1999.0634 URL [本文引用: 1]
Notes on extra-European taxa of Bolbitiaceae (Agaricales, Basidiomycota)
Colored illustrations of macrofungi (mushrooms) of China
Long-term follow-up of psilocybin-facilitated smoking cessation
DOI:10.3109/00952990.2016.1170135 URL [本文引用: 1]
Checklist of the genera Conocybe and Pholiotina (Agaricales, Agaricomycetes) in Estonia
DOI:10.12697/fce.2013.50.05 URL [本文引用: 1]
ModelFinder: fast model selection for accurate phylogenetic estimates
DOI:10.1038/nmeth.4285
PMID:28481363
[本文引用: 1]
Model-based molecular phylogenetics plays an important role in comparisons of genomic data, and model selection is a key step in all such analyses. We present ModelFinder, a fast model-selection method that greatly improves the accuracy of phylogenetic estimates by incorporating a model of rate heterogeneity across sites not previously considered in this context and by allowing concurrent searches of model space and tree space.
Morphological observation and molecular identification of macrofungi in Baluntai Steppe
Macrofungus flora of Hunnan
Mushrooms of Changbai Mountains, China
Atlas of Chinese macrofungal resources
Taxonomy and molecular phylogeny of Bolbitiaceae in Northeast China
New species and new records in the genus Conocybe (Bolbitaceae) from China
DOI:10.11646/phytotaxa.357.4 URL [本文引用: 2]
Economic fungi of China
The macrofungi in China
Economic macrofungi of Tibet
Fungi of the Qinling Mountains
Major clades of Agaricales: a multilocus phylogenetic overview
An overview of the phylogeny of the Agaricales is presented based on a multilocus analysis of a six-gene region supermatrix. Bayesian analyses of 5611 nucleotide characters of rpb1, rpb1-intron 2, rpb2 and 18S, 25S, and 5.8S ribosomal RNA genes recovered six major clades, which are recognized informally and labeled the Agaricoid, Tricholomatoid, Marasmioid, Pluteoid, Hygrophoroid and Plicaturopsidoid clades. Each clade is discussed in terms of key morphological and ecological traits. At least 11 origins of the ectomycorrhizal habit appear to have evolved in the Agaricales, with possibly as many as nine origins in the Agaricoid plus Tricholomatoid clade alone. A family-based phylogenetic classification is sketched for the Agaricales, in which 30 families, four unplaced tribes and two informally named clades are recognized.
Ultrafast approximation for phylogenetic bootstrap
DOI:10.1093/molbev/mst024
PMID:23418397
[本文引用: 1]
Nonparametric bootstrap has been a widely used tool in phylogenetic analysis to assess the clade support of phylogenetic trees. However, with the rapidly growing amount of data, this task remains a computational bottleneck. Recently, approximation methods such as the RAxML rapid bootstrap (RBS) and the Shimodaira-Hasegawa-like approximate likelihood ratio test have been introduced to speed up the bootstrap. Here, we suggest an ultrafast bootstrap approximation approach (UFBoot) to compute the support of phylogenetic groups in maximum likelihood (ML) based trees. To achieve this, we combine the resampling estimated log-likelihood method with a simple but effective collection scheme of candidate trees. We also propose a stopping rule that assesses the convergence of branch support values to automatically determine when to stop collecting candidate trees. UFBoot achieves a median speed up of 3.1 (range: 0.66-33.3) to 10.2 (range: 1.32-41.4) compared with RAxML RBS for real DNA and amino acid alignments, respectively. Moreover, our extensive simulations show that UFBoot is robust against moderate model violations and the support values obtained appear to be relatively unbiased compared with the conservative standard bootstrap. This provides a more direct interpretation of the bootstrap support. We offer an efficient and easy-to-use software (available at http://www.cibiv.at/software/iqtree) to perform the UFBoot analysis with ML tree inference.
Phylogenetic relationships of agaric fungi based on nuclear large subunit ribosomal DNA sequences
DOI:10.1093/sysbio/49.2.278
PMID:12118409
[本文引用: 1]
Phylogenetic relationships of mushrooms and their relatives within the order Agaricales were addressed by using nuclear large subunit ribosomal DNA sequences. Approximately 900 bases of the 5' end of the nucleus-encoded large subunit RNA gene were sequenced for 154 selected taxa representing most families within the Agaricales. Several phylogenetic methods were used, including weighted and equally weighted parsimony (MP), maximum likelihood (ML), and distance methods (NJ). The starting tree for branch swapping in the ML analyses was the tree with the highest ML score among previously produced MP and NJ trees. A high degree of consensus was observed between phylogenetic estimates obtained through MP and ML. NJ trees differed according to the distance model that was used; however, all NJ trees still supported most of the same terminal groupings as the MP and ML trees did. NJ trees were always significantly suboptimal when evaluated against the best MP and ML trees, by both parsimony and likelihood tests. Our analyses suggest that weighted MP and ML provide the best estimates of Agaricales phylogeny. Similar support was observed between bootstrapping and jackknifing methods for evaluation of tree robustness. Phylogenetic analyses revealed many groups of agaricoid fungi that are supported by moderate to high bootstrap or jackknife values or are consistent with morphology-based classification schemes. Analyses also support separate placement of the boletes and russules, which are basal to the main core group of gilled mushrooms (the Agaricineae of Singer). Examples of monophyletic groups include the families Amanitaceae, Coprinaceae (excluding Coprinus comatus and subfamily Panaeolideae), Agaricaceae (excluding the Cystodermateae), and Strophariaceae pro parte (Stropharia, Pholiota, and Hypholoma); the mycorrhizal species of Tricholoma (including Leucopaxillus, also mycorrhizal); Mycena and Resinomycena; Termitomyces, Podabrella, and Lyophyllum; and Pleurotus with Hohenbuehelia. Several groups revealed by these data to be nonmonophyletic include the families Tricholomataceae, Cortinariaceae, and Hygrophoraceae and the genera Clitocybe, Omphalina, and Marasmius. This study provides a framework for future systematics studies in the Agaricales and suggestions for analyzing large molecular data sets.
One hundred and seventeen clades of euagarics
DOI:10.1016/S1055-7903(02)00027-1 URL [本文引用: 1]
Asproinocybaceae fam. nov.(Agaricales, Agaricomycetes) for accommodating the genera Asproinocybe and Tricholosporum, and description of Asproinocybe sinensis and Tricholosporum guangxiense sp. nov
DOI:10.3390/jof7121086
URL
[本文引用: 1]
Asproinocybe and Tricholosporum are not well known, and their placement at the family level remains undetermined. In this study, we conducted molecular phylogenetic analyses based on nuc rDNA internal transcribed spacer region (ITS) and nuc 28S rDNA (nrLSU), and a dataset with six molecular markers (ITS, LSU, RNA polymerase II largest subunit (RPB1), RNA polymerase II second largest subunit (RPB2), 18S nuclear small subunit ribosomal DNA (nrSSU), and translation elongation factor 1-alpha (TEF1-α)) using Bayesian (BA) and Maximum Likelihood (ML) methods, we found that the species of Asproinocybe and Tricholosporum formed an independent family-level clade (0.98/72). Asproinocybaceae fam. nov., a new family, is established here for accommodating this clade. Two new species, Asproinocybe sinensis and Tricholosporum guangxiense, from subtropical and tropical karst areas of China, are also described here.
IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies
DOI:10.1093/molbev/msu300 URL [本文引用: 1]
How ecstasy and psilocybin are shaking up psychiatry
DOI:10.1038/d41586-021-00187-9 [本文引用: 1]
MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space
DOI:10.1093/sysbio/sys029
PMID:22357727
[本文引用: 1]
Since its introduction in 2001, MrBayes has grown in popularity as a software package for Bayesian phylogenetic inference using Markov chain Monte Carlo (MCMC) methods. With this note, we announce the release of version 3.2, a major upgrade to the latest official release presented in 2003. The new version provides convergence diagnostics and allows multiple analyses to be run in parallel with convergence progress monitored on the fly. The introduction of new proposals and automatic optimization of tuning parameters has improved convergence for many problems. The new version also sports significantly faster likelihood calculations through streaming single-instruction-multiple-data extensions (SSE) and support of the BEAGLE library, allowing likelihood calculations to be delegated to graphics processing units (GPUs) on compatible hardware. Speedup factors range from around 2 with SSE code to more than 50 with BEAGLE for codon problems. Checkpointing across all models allows long runs to be completed even when an analysis is prematurely terminated. New models include relaxed clocks, dating, model averaging across time-reversible substitution models, and support for hard, negative, and partial (backbone) tree constraints. Inference of species trees from gene trees is supported by full incorporation of the Bayesian estimation of species trees (BEST) algorithms. Marginal model likelihoods for Bayes factor tests can be estimated accurately across the entire model space using the stepping stone method. The new version provides more output options than previously, including samples of ancestral states, site rates, site d(N)/d(S) rations, branch rates, and node dates. A wide range of statistics on tree parameters can also be output for visualization in FigTree and compatible software.
Forest mushroom in China
New and interesting species of Basidiomycetes
New taxa and new combinations of Agaricales (Diagnoses fungorum novorum Agaricalium 4)
Notes on Conocybe (Bolbitiaceae)
DOI:10.1007/BF00937429 URL [本文引用: 2]
The group of Conocybe mesospora in Europe (Bolbitiaceae)
DOI:10.1007/BF00940399 URL [本文引用: 1]
Sylloge fungorum sinicorum
Iteratively refined guide trees help improving alignment and phylogenetic inference in the mushroom family Bolbitiaceae
DOI:10.1371/journal.pone.0056143 URL [本文引用: 4]
The genus Conocybe subgen. Pholiotina. I. The European annulate species
Novitates mycologicae novissimae
Research on the diversity of macrofungi on campus of Hanjiang Normal University
Biodiversity of macrofungi in Changbai Mountain region
Observations on the Bolbitiaceae-IV. Developmental studies on Conocybe with particular reference to the annulate species
Observations on the Bolbitiaceae. 16. Conocybe sect. Gigantae
British fungus flora. Agarics and boleti 3. Bolbitiaceae: Agrocybe, Bolbitius and Conocybe
Observations on the Bolbitiaceae-23. Interesting Danish members of the family
DOI:10.1111/j.1756-1051.1983.tb01077.x URL [本文引用: 1]
Observations on the Bolbitiaceae. 26. Bolbitiaceae of Kashmir with particular reference to the genus Agrocybe
Observations on the Bolbitiaceae 31
Observations on the Bolbitiaceae: 30 Agaricus callistus Peck
Resource diversity of Chinese macrofungi: edible, medicinal and poisonous species
DOI:10.1007/s13225-019-00432-7 [本文引用: 1]
Fungi of tropical China
Illustrations of agarics of Changbai Mountains, China
Studies on macrofungi diversity in Jianshan Nature Reserve, Wenxian County
Mushrooms of Sichuan
The pictorial book of mushrooms of China
Fungi of the Hengduan Mountains
PhyloSuite: an integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies
DOI:10.1111/1755-0998.13096
PMID:31599058
[本文引用: 1]
Multigene and genomic data sets have become commonplace in the field of phylogenetics, but many existing tools are not designed for such data sets, which often makes the analysis time-consuming and tedious. Here, we present PhyloSuite, a (cross-platform, open-source, stand-alone Python graphical user interface) user-friendly workflow desktop platform dedicated to streamlining molecular sequence data management and evolutionary phylogenetics studies. It uses a plugin-based system that integrates several phylogenetic and bioinformatic tools, thereby streamlining the entire procedure, from data acquisition to phylogenetic tree annotation (in combination with iTOL). It has the following features: (a) point-and-click and drag-and-drop graphical user interface; (b) a workplace to manage and organize molecular sequence data and results of analyses; (c) GenBank entry extraction and comparative statistics; and (d) a phylogenetic workflow with batch processing capability, comprising sequence alignment (mafft and macse), alignment optimization (trimAl, HmmCleaner and Gblocks), data set concatenation, best partitioning scheme and best evolutionary model selection (PartitionFinder and modelfinder), and phylogenetic inference (MrBayes and iq-tree). PhyloSuite is designed for both beginners and experienced researchers, allowing the former to quick-start their way into phylogenetic analysis, and the latter to conduct, store and manage their work in a streamlined way, and spend more time investigating scientific questions instead of wasting it on transferring files from one software program to another.© 2019 John Wiley & Sons Ltd.
Hong Kong mushrooms
Investigation of macrofungi resources in Shennongjia and artificial culture of Cordyceps cicadae fruit body
Taxonomy of coprinoid fungi in China
中国森林大型真菌重要类群多样性和系统学研究



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}