 PDF(2453 KB)
PDF(2453 KB)

皱皮粗柄白蚁伞子实体和蚁巢的关联物质分析及氨基酸糖类促生作用初探
王成凤, 刘夏, 张曦予, 董杰, 张实润, 杨晓敏, 吕为, 刘佳艳, 李宗菊
菌物学报 ›› 2025, Vol. 44 ›› Issue (3) : 240256.
PDF(2453 KB)
PDF(2453 KB)
皱皮粗柄白蚁伞子实体和蚁巢的关联物质分析及氨基酸糖类促生作用初探
 ({{custom_author.role_cn}}), {{javascript:window.custom_author_cn_index++;}}
({{custom_author.role_cn}}), {{javascript:window.custom_author_cn_index++;}}Analyses of substances associating Termitomyces robustus fruiting bodies with termite nests and preliminary studies on growth-promoting effect of amino acids and sugars
({{custom_author.role_en}}), {{javascript:window.custom_author_en_index++;}}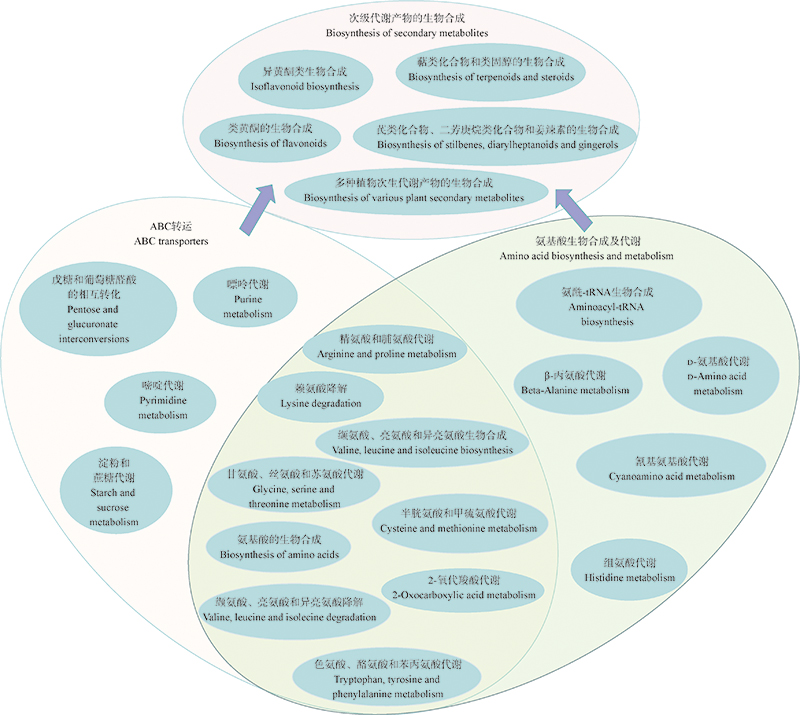
| {{custom_ref.label}} |
{{custom_citation.content}}
{{custom_citation.annotation}}
|


/
| 〈 |
|
〉 |







