PDF(1165 KB)
PDF(1165 KB)

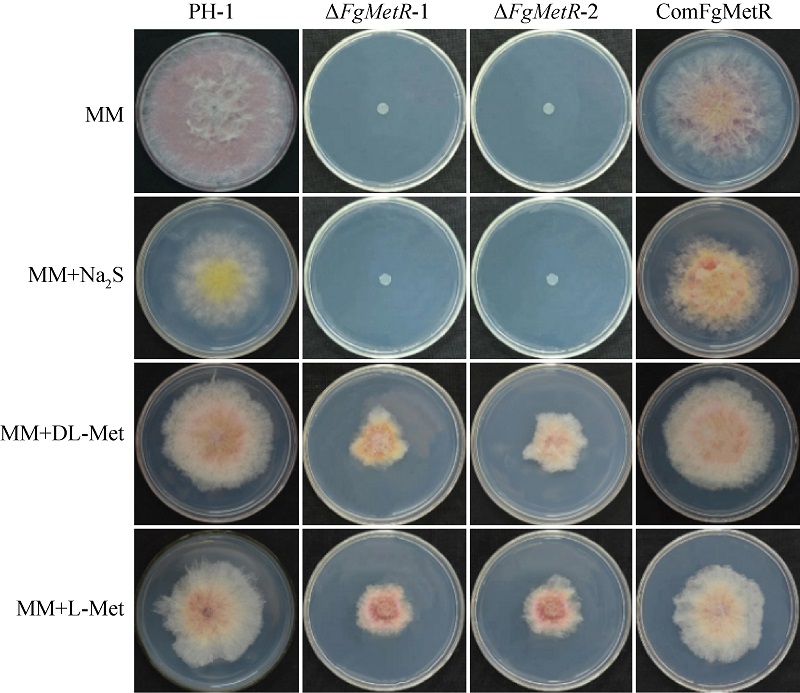
Biological functions of the bZIP transcription factor FgMetR in Fusarium graminearum
MA Hao, ZHANG Limin, ZHAO Yanxiang, HUANG Jinguang
Mycosystema ›› 2025, Vol. 44 ›› Issue (4) : 240279.
PDF(1165 KB)
PDF(1165 KB)
Biological functions of the bZIP transcription factor FgMetR in Fusarium graminearum
 ({{custom_author.role_en}})
,
{{javascript:window.custom_author_en_index++;}}
({{custom_author.role_en}})
,
{{javascript:window.custom_author_en_index++;}}
| {{custom_ref.label}} |
{{custom_citation.content}}
{{custom_citation.annotation}}
|


/
| 〈 |
|
〉 |