PDF(1422 KB)
PDF(1422 KB)

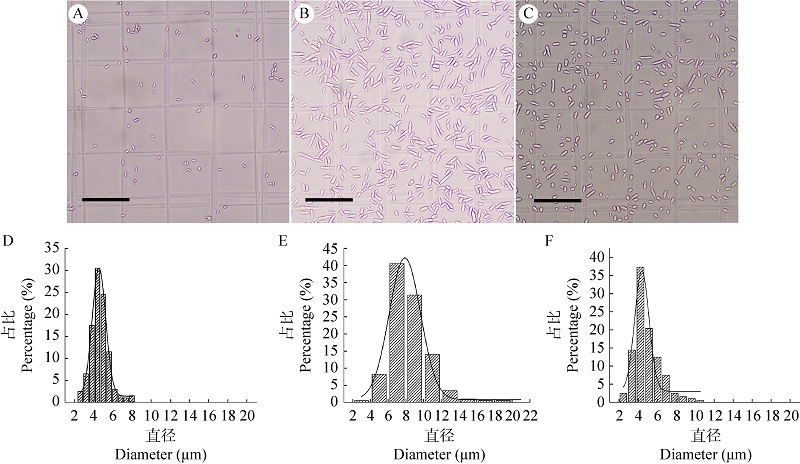
Surface characteristics and stress tolerance of submerged conidia, blastospores and aerial conidia of biocontrol fungus Cordyceps javanica IF-1106
GAO Meiyu, LI Junmei, LI Yihua, XIANG Huiming, MA Ruiyan, ZHOU Wenwen
Mycosystema ›› 2025, Vol. 44 ›› Issue (4) : 240288.
PDF(1422 KB)
PDF(1422 KB)
Surface characteristics and stress tolerance of submerged conidia, blastospores and aerial conidia of biocontrol fungus Cordyceps javanica IF-1106
 ({{custom_author.role_en}})
,
{{javascript:window.custom_author_en_index++;}}
({{custom_author.role_en}})
,
{{javascript:window.custom_author_en_index++;}}
| {{custom_ref.label}} |
{{custom_citation.content}}
{{custom_citation.annotation}}
|


/
| 〈 |
|
〉 |