Population genetic structure of Colletotrichum fructicola
ZHU Dan-Xue1, 2, ZHOU Guo-Ying1, XU Jian-Ping1, LIU Jun-Ang1, LI He1, *,
1Key Laboratory for Non-wood Forest Cultivation and Conservation of Ministry of Education, Central South University of Forestry and Technology, Changsha, Hunan 410004, China2College of Life Science and Technology, Central South University of Forestry and Technology, Changsha, Hunan 410004, China
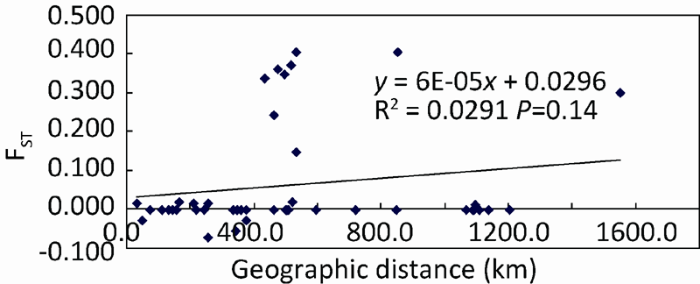
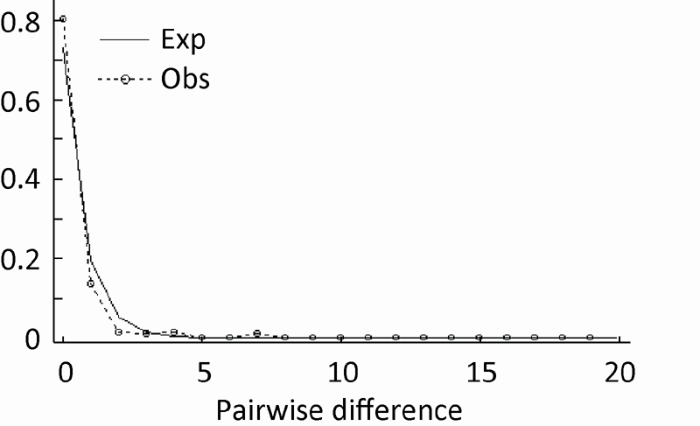
nrDNA ITS sequences were used to analyze the genetic structure and genetic diversity of Colletotrichum fructicola populations consisting of 145 strains isolated from 10 sites. A total of 13 ITS haplotypes were identified among which the dominant haplotype (haplotype 5) included 127 specimens from the 10 sites. The genetic differentiation index, FST, revealed significant genetic differentiation between Suizhou populations and the others. AMOVA test showed that 13% of genetic differentiation occurred among population and 87% within population. The Mantel test showed no linear-correlation between genetic distance and geographical distance. Mismatch distribution analysis suggested there was population expansion for Colletotrichum fructicola and gene flow was detected between populations.
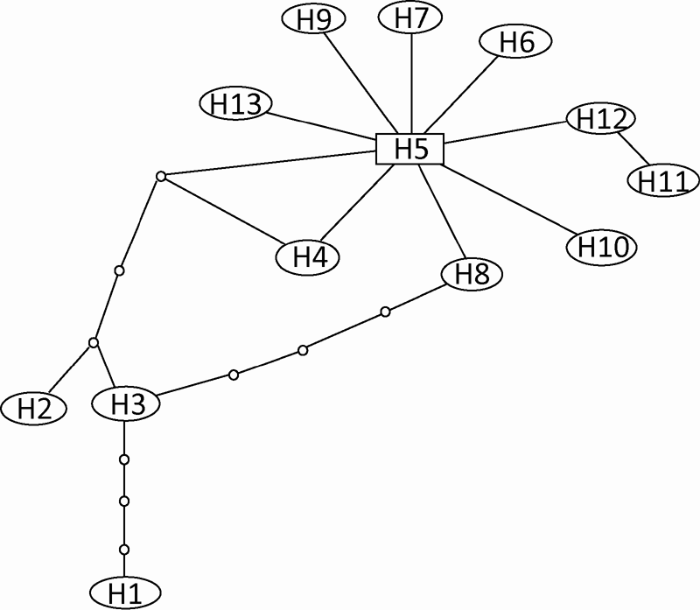
Fig. 1 Parsimony network of the haplotypes of Colletotrichum fructicola. Each mutation is shown as a line between each two haplotypes. Small dots represent unsampled haplotypes.
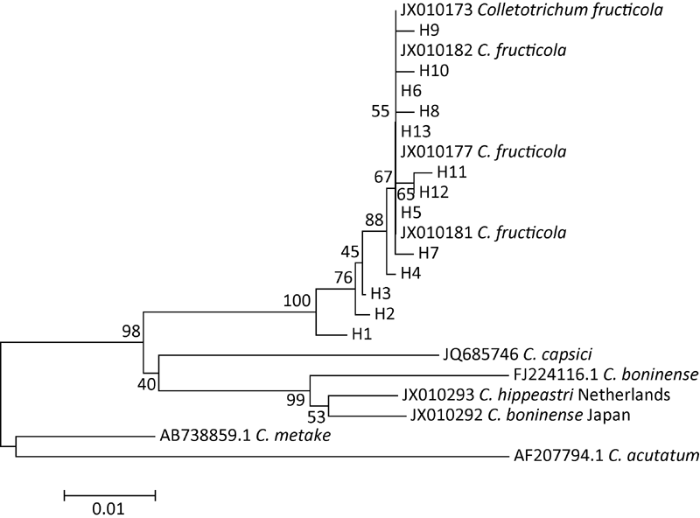
Fig. 4 A neighbour-joining tree of the ITS sequences.
3 讨论
核糖体DNA内转录间隔区ITS(internal transcribed spacer)(包括5.8S)是介于18S和26S之间的非编码转录间隔区,承受的选择压力较小,进化速率快,能提供比较丰富的核苷酸变异位点和信息位点,且由于协同进化使得这个片段在不同的重复单元之间十分一致,此外ITS序列片段大小适中,易于测序和分析,近年来被广泛地应用于解决较低分类单元层次上的真菌系统发育问题(Seifert 2009;Bellemain et al. 2010)。Sha et al.(2008)用ITS技术研究云南干巴菌Thelephora ganbajun的遗传多样性,利用通用引物ITS4和ITS5扩增该菌156个个体的ITS区域,结果表明不同地理来源的种群间变异为0.825,种群内变异为0.175,即该菌遗传分化较大。刘诗胤等(2012)利用ITS测序技术研究了浙江地区不同寄主来源炭疽菌的遗传多样性,结果表明不同菌株间的ITS具有丰富的多态性。刘芳等(2011)应用rDNA-ITS序列分析方法研究悬钩子皮下盘菌种内的遗传多样性,序列比较分析表明,悬钩子皮下盘菌ITS序列长度为443-456bp,G+C含量为48.7%-51.5%;不同菌株间的ITS区序列变异较大,该菌种内遗传多样性丰富。张振等(2009)对陕西省31 株木本植物炭疽菌的rDNA-ITS区序列进行了分析,发现炭疽菌rDNA的ITS区序列存在明显的差异,其ITS-rDNA序列多样性达0.1957,序列长度差异达94bp,通过聚类分析可将其分为不同的遗传类群。因此,炭疽菌rDNA的ITS区序列对种及种下专化型的划分具有一定的意义。本研究利用ITS序列,分析了我国果生刺盘孢菌的种群遗传结构,发现该病菌在我国油茶产区分布十分广泛,单倍型多样性较丰富,存在一定的遗传分化现象。Kelemu et al.(1997)对45个来自南美的圭亚那柱花草盘长孢状刺盘孢菌Colletotrichum gloeosporioides Penz.进行了RAPD遗传多态性分析,结果表明柱花草盘长孢状刺盘孢菌具有显著多态性,来自长期种植区的菌株的遗传多态性比新植区的丰富,这与本研究中,湖南天际岭和湖北随州这些超过50年树龄的地区病菌单倍型多样性丰富度一致。
CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice.
Progress, problem and prospect of oil camelliae anthracnose (Colletotrichum gloeosporioides) research.
Forest Pest and Disease, 28(2): 27-31 (in Chinese)
[9]
KelemuS, BadelJL, MorenoCX, 1997.
Biodiversity, epidemiology and virulence of Colletotrichum gloeosporioides. I. Genetic and pathogenic diversity in Colletotrichum gloeosporioides isolates from Stylosanthes guianensis.
WrightS, 1978. Evolution and the genetics of populations. Volume 4. Variability with in and among natural populations. The University of Chicago Press, Chicago. 325-327
ITS as an environmental DNA barcode for fungi: an insilico approach reveals potential PCR biases.
1
2010
... 核糖体DNA内转录间隔区ITS(internal transcribed spacer)(包括5.8S)是介于18S和26S之间的非编码转录间隔区,承受的选择压力较小,进化速率快,能提供比较丰富的核苷酸变异位点和信息位点,且由于协同进化使得这个片段在不同的重复单元之间十分一致,此外ITS序列片段大小适中,易于测序和分析,近年来被广泛地应用于解决较低分类单元层次上的真菌系统发育问题(Seifert 2009;Bellemain et al. 2010).Sha et al.(2008)用ITS技术研究云南干巴菌Thelephora ganbajun的遗传多样性,利用通用引物ITS4和ITS5扩增该菌156个个体的ITS区域,结果表明不同地理来源的种群间变异为0.825,种群内变异为0.175,即该菌遗传分化较大.刘诗胤等(2012)利用ITS测序技术研究了浙江地区不同寄主来源炭疽菌的遗传多样性,结果表明不同菌株间的ITS具有丰富的多态性.刘芳等(2011)应用rDNA-ITS序列分析方法研究悬钩子皮下盘菌种内的遗传多样性,序列比较分析表明,悬钩子皮下盘菌ITS序列长度为443-456bp,G+C含量为48.7%-51.5%;不同菌株间的ITS区序列变异较大,该菌种内遗传多样性丰富.张振等(2009)对陕西省31 株木本植物炭疽菌的rDNA-ITS区序列进行了分析,发现炭疽菌rDNA的ITS区序列存在明显的差异,其ITS-rDNA序列多样性达0.1957,序列长度差异达94bp,通过聚类分析可将其分为不同的遗传类群.因此,炭疽菌rDNA的ITS区序列对种及种下专化型的划分具有一定的意义.本研究利用ITS序列,分析了我国果生刺盘孢菌的种群遗传结构,发现该病菌在我国油茶产区分布十分广泛,单倍型多样性较丰富,存在一定的遗传分化现象.Kelemu et al.(1997)对45个来自南美的圭亚那柱花草盘长孢状刺盘孢菌Colletotrichum gloeosporioides Penz.进行了RAPD遗传多态性分析,结果表明柱花草盘长孢状刺盘孢菌具有显著多态性,来自长期种植区的菌株的遗传多态性比新植区的丰富,这与本研究中,湖南天际岭和湖北随州这些超过50年树龄的地区病菌单倍型多样性丰富度一致. ...
A software tool for simple and partial Mantel tests.
1
2002
... 采用BioEdit软件(Hall 1999)对DNA序列进行比对和编辑,同时辅以手工矫正.用Mega 5.0(Tamura et al. 2011)软件统计碱基组成、变异位点数等序列特征.使用GenALEx6(Peakall & Smouse 2006)分析各不同地区果生刺盘孢菌群体的ITS单倍型,研究:(1)各不同群体的单倍型多样性;(2)不同地区群体之间的遗传关系;(3)遗传分化与地理隔离的关系.其中,每个单独的群体及群体间的单倍型多样性和群体遗传分化FST值分开计算.遗传分化与地理距离的关系使用Mantel 检验法分析(Bonnet & Vander Peer 2002).地理群体间的距离使用经纬度与坐标共同表示.最后,分子变异分析(analysis of molecular variance,AMOVA)用于估算地理隔离对病菌ITS单倍型分布的相对影响(Excoffier et al. 1992).用DnaSP 5.0软件(Librado & Rozas 2009)统计ITS序列的核苷酸多样性(nucleotide diversity,Pi)和群体间遗传分化指数(fixation index,FST);根据ITS序列的单倍型频率和核苷酸差异,用 TCS1.21软件(Clement et al. 2000)构建单倍型的网状亲缘关系图.采用DnaSP5.0软件对所有地理种群序列的单倍型错配进行分析.第二种类型的分析中,每个ITS单倍型序列都用于从GenBank中搜索近缘的序列.然后用Clustal W软件(Higgins et al. 1994)对我们的序列及搜索得到的序列进行排列,手工适当校正后用Mega 5.0软件进行分析.使用 Kimura双参数模型,排列中产生的空位(gap)被处理为缺失(missing)构建分子系统发育树(N-J tree),自展法(Bootstrap,1 000次重复)检验各分支的置信度(Tamura et al. 2004). ...
TCS: a computer program to estimate gene genealogies.
1
2000
... 采用BioEdit软件(Hall 1999)对DNA序列进行比对和编辑,同时辅以手工矫正.用Mega 5.0(Tamura et al. 2011)软件统计碱基组成、变异位点数等序列特征.使用GenALEx6(Peakall & Smouse 2006)分析各不同地区果生刺盘孢菌群体的ITS单倍型,研究:(1)各不同群体的单倍型多样性;(2)不同地区群体之间的遗传关系;(3)遗传分化与地理隔离的关系.其中,每个单独的群体及群体间的单倍型多样性和群体遗传分化FST值分开计算.遗传分化与地理距离的关系使用Mantel 检验法分析(Bonnet & Vander Peer 2002).地理群体间的距离使用经纬度与坐标共同表示.最后,分子变异分析(analysis of molecular variance,AMOVA)用于估算地理隔离对病菌ITS单倍型分布的相对影响(Excoffier et al. 1992).用DnaSP 5.0软件(Librado & Rozas 2009)统计ITS序列的核苷酸多样性(nucleotide diversity,Pi)和群体间遗传分化指数(fixation index,FST);根据ITS序列的单倍型频率和核苷酸差异,用 TCS1.21软件(Clement et al. 2000)构建单倍型的网状亲缘关系图.采用DnaSP5.0软件对所有地理种群序列的单倍型错配进行分析.第二种类型的分析中,每个ITS单倍型序列都用于从GenBank中搜索近缘的序列.然后用Clustal W软件(Higgins et al. 1994)对我们的序列及搜索得到的序列进行排列,手工适当校正后用Mega 5.0软件进行分析.使用 Kimura双参数模型,排列中产生的空位(gap)被处理为缺失(missing)构建分子系统发育树(N-J tree),自展法(Bootstrap,1 000次重复)检验各分支的置信度(Tamura et al. 2004). ...
Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data.
1
1992
... 采用BioEdit软件(Hall 1999)对DNA序列进行比对和编辑,同时辅以手工矫正.用Mega 5.0(Tamura et al. 2011)软件统计碱基组成、变异位点数等序列特征.使用GenALEx6(Peakall & Smouse 2006)分析各不同地区果生刺盘孢菌群体的ITS单倍型,研究:(1)各不同群体的单倍型多样性;(2)不同地区群体之间的遗传关系;(3)遗传分化与地理隔离的关系.其中,每个单独的群体及群体间的单倍型多样性和群体遗传分化FST值分开计算.遗传分化与地理距离的关系使用Mantel 检验法分析(Bonnet & Vander Peer 2002).地理群体间的距离使用经纬度与坐标共同表示.最后,分子变异分析(analysis of molecular variance,AMOVA)用于估算地理隔离对病菌ITS单倍型分布的相对影响(Excoffier et al. 1992).用DnaSP 5.0软件(Librado & Rozas 2009)统计ITS序列的核苷酸多样性(nucleotide diversity,Pi)和群体间遗传分化指数(fixation index,FST);根据ITS序列的单倍型频率和核苷酸差异,用 TCS1.21软件(Clement et al. 2000)构建单倍型的网状亲缘关系图.采用DnaSP5.0软件对所有地理种群序列的单倍型错配进行分析.第二种类型的分析中,每个ITS单倍型序列都用于从GenBank中搜索近缘的序列.然后用Clustal W软件(Higgins et al. 1994)对我们的序列及搜索得到的序列进行排列,手工适当校正后用Mega 5.0软件进行分析.使用 Kimura双参数模型,排列中产生的空位(gap)被处理为缺失(missing)构建分子系统发育树(N-J tree),自展法(Bootstrap,1 000次重复)检验各分支的置信度(Tamura et al. 2004). ...
Genetic diversity of Lactarius deliciosus from South China.
0
2011
BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT.
1
1999
... 采用BioEdit软件(Hall 1999)对DNA序列进行比对和编辑,同时辅以手工矫正.用Mega 5.0(Tamura et al. 2011)软件统计碱基组成、变异位点数等序列特征.使用GenALEx6(Peakall & Smouse 2006)分析各不同地区果生刺盘孢菌群体的ITS单倍型,研究:(1)各不同群体的单倍型多样性;(2)不同地区群体之间的遗传关系;(3)遗传分化与地理隔离的关系.其中,每个单独的群体及群体间的单倍型多样性和群体遗传分化FST值分开计算.遗传分化与地理距离的关系使用Mantel 检验法分析(Bonnet & Vander Peer 2002).地理群体间的距离使用经纬度与坐标共同表示.最后,分子变异分析(analysis of molecular variance,AMOVA)用于估算地理隔离对病菌ITS单倍型分布的相对影响(Excoffier et al. 1992).用DnaSP 5.0软件(Librado & Rozas 2009)统计ITS序列的核苷酸多样性(nucleotide diversity,Pi)和群体间遗传分化指数(fixation index,FST);根据ITS序列的单倍型频率和核苷酸差异,用 TCS1.21软件(Clement et al. 2000)构建单倍型的网状亲缘关系图.采用DnaSP5.0软件对所有地理种群序列的单倍型错配进行分析.第二种类型的分析中,每个ITS单倍型序列都用于从GenBank中搜索近缘的序列.然后用Clustal W软件(Higgins et al. 1994)对我们的序列及搜索得到的序列进行排列,手工适当校正后用Mega 5.0软件进行分析.使用 Kimura双参数模型,排列中产生的空位(gap)被处理为缺失(missing)构建分子系统发育树(N-J tree),自展法(Bootstrap,1 000次重复)检验各分支的置信度(Tamura et al. 2004). ...
CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice.
1
1994
... 采用BioEdit软件(Hall 1999)对DNA序列进行比对和编辑,同时辅以手工矫正.用Mega 5.0(Tamura et al. 2011)软件统计碱基组成、变异位点数等序列特征.使用GenALEx6(Peakall & Smouse 2006)分析各不同地区果生刺盘孢菌群体的ITS单倍型,研究:(1)各不同群体的单倍型多样性;(2)不同地区群体之间的遗传关系;(3)遗传分化与地理隔离的关系.其中,每个单独的群体及群体间的单倍型多样性和群体遗传分化FST值分开计算.遗传分化与地理距离的关系使用Mantel 检验法分析(Bonnet & Vander Peer 2002).地理群体间的距离使用经纬度与坐标共同表示.最后,分子变异分析(analysis of molecular variance,AMOVA)用于估算地理隔离对病菌ITS单倍型分布的相对影响(Excoffier et al. 1992).用DnaSP 5.0软件(Librado & Rozas 2009)统计ITS序列的核苷酸多样性(nucleotide diversity,Pi)和群体间遗传分化指数(fixation index,FST);根据ITS序列的单倍型频率和核苷酸差异,用 TCS1.21软件(Clement et al. 2000)构建单倍型的网状亲缘关系图.采用DnaSP5.0软件对所有地理种群序列的单倍型错配进行分析.第二种类型的分析中,每个ITS单倍型序列都用于从GenBank中搜索近缘的序列.然后用Clustal W软件(Higgins et al. 1994)对我们的序列及搜索得到的序列进行排列,手工适当校正后用Mega 5.0软件进行分析.使用 Kimura双参数模型,排列中产生的空位(gap)被处理为缺失(missing)构建分子系统发育树(N-J tree),自展法(Bootstrap,1 000次重复)检验各分支的置信度(Tamura et al. 2004). ...
Progress, problem and prospect of oil camelliae anthracnose (Colletotrichum gloeosporioides) research.
0
2009
Biodiversity, epidemiology and virulence of Colletotrichum gloeosporioides. I. Genetic and pathogenic diversity in Colletotrichum gloeosporioides isolates from Stylosanthes guianensis.
1
1997
... 核糖体DNA内转录间隔区ITS(internal transcribed spacer)(包括5.8S)是介于18S和26S之间的非编码转录间隔区,承受的选择压力较小,进化速率快,能提供比较丰富的核苷酸变异位点和信息位点,且由于协同进化使得这个片段在不同的重复单元之间十分一致,此外ITS序列片段大小适中,易于测序和分析,近年来被广泛地应用于解决较低分类单元层次上的真菌系统发育问题(Seifert 2009;Bellemain et al. 2010).Sha et al.(2008)用ITS技术研究云南干巴菌Thelephora ganbajun的遗传多样性,利用通用引物ITS4和ITS5扩增该菌156个个体的ITS区域,结果表明不同地理来源的种群间变异为0.825,种群内变异为0.175,即该菌遗传分化较大.刘诗胤等(2012)利用ITS测序技术研究了浙江地区不同寄主来源炭疽菌的遗传多样性,结果表明不同菌株间的ITS具有丰富的多态性.刘芳等(2011)应用rDNA-ITS序列分析方法研究悬钩子皮下盘菌种内的遗传多样性,序列比较分析表明,悬钩子皮下盘菌ITS序列长度为443-456bp,G+C含量为48.7%-51.5%;不同菌株间的ITS区序列变异较大,该菌种内遗传多样性丰富.张振等(2009)对陕西省31 株木本植物炭疽菌的rDNA-ITS区序列进行了分析,发现炭疽菌rDNA的ITS区序列存在明显的差异,其ITS-rDNA序列多样性达0.1957,序列长度差异达94bp,通过聚类分析可将其分为不同的遗传类群.因此,炭疽菌rDNA的ITS区序列对种及种下专化型的划分具有一定的意义.本研究利用ITS序列,分析了我国果生刺盘孢菌的种群遗传结构,发现该病菌在我国油茶产区分布十分广泛,单倍型多样性较丰富,存在一定的遗传分化现象.Kelemu et al.(1997)对45个来自南美的圭亚那柱花草盘长孢状刺盘孢菌Colletotrichum gloeosporioides Penz.进行了RAPD遗传多态性分析,结果表明柱花草盘长孢状刺盘孢菌具有显著多态性,来自长期种植区的菌株的遗传多态性比新植区的丰富,这与本研究中,湖南天际岭和湖北随州这些超过50年树龄的地区病菌单倍型多样性丰富度一致. ...
DnaSP v5: a software for comprehensive analysis of DNA polymorphism data.
1
2009
... 采用BioEdit软件(Hall 1999)对DNA序列进行比对和编辑,同时辅以手工矫正.用Mega 5.0(Tamura et al. 2011)软件统计碱基组成、变异位点数等序列特征.使用GenALEx6(Peakall & Smouse 2006)分析各不同地区果生刺盘孢菌群体的ITS单倍型,研究:(1)各不同群体的单倍型多样性;(2)不同地区群体之间的遗传关系;(3)遗传分化与地理隔离的关系.其中,每个单独的群体及群体间的单倍型多样性和群体遗传分化FST值分开计算.遗传分化与地理距离的关系使用Mantel 检验法分析(Bonnet & Vander Peer 2002).地理群体间的距离使用经纬度与坐标共同表示.最后,分子变异分析(analysis of molecular variance,AMOVA)用于估算地理隔离对病菌ITS单倍型分布的相对影响(Excoffier et al. 1992).用DnaSP 5.0软件(Librado & Rozas 2009)统计ITS序列的核苷酸多样性(nucleotide diversity,Pi)和群体间遗传分化指数(fixation index,FST);根据ITS序列的单倍型频率和核苷酸差异,用 TCS1.21软件(Clement et al. 2000)构建单倍型的网状亲缘关系图.采用DnaSP5.0软件对所有地理种群序列的单倍型错配进行分析.第二种类型的分析中,每个ITS单倍型序列都用于从GenBank中搜索近缘的序列.然后用Clustal W软件(Higgins et al. 1994)对我们的序列及搜索得到的序列进行排列,手工适当校正后用Mega 5.0软件进行分析.使用 Kimura双参数模型,排列中产生的空位(gap)被处理为缺失(missing)构建分子系统发育树(N-J tree),自展法(Bootstrap,1 000次重复)检验各分支的置信度(Tamura et al. 2004). ...
ITS sequence analysis of intraspecific genetic diversity of Hypoderma rubi.
0
2011
RAPD analysis of the pathogenic fungi of anthracnose of fruits.
0
2002
Polymorphism analysis of Colletotrichum spp. from Zhejiang Province by ITS rDNA PCR-RFLP.
0
2012
Studies on etiology, occurrence and control of camellia oleifera anthracnose. Master Thesis,
0
2012
GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research.
1
2006
... 采用BioEdit软件(Hall 1999)对DNA序列进行比对和编辑,同时辅以手工矫正.用Mega 5.0(Tamura et al. 2011)软件统计碱基组成、变异位点数等序列特征.使用GenALEx6(Peakall & Smouse 2006)分析各不同地区果生刺盘孢菌群体的ITS单倍型,研究:(1)各不同群体的单倍型多样性;(2)不同地区群体之间的遗传关系;(3)遗传分化与地理隔离的关系.其中,每个单独的群体及群体间的单倍型多样性和群体遗传分化FST值分开计算.遗传分化与地理距离的关系使用Mantel 检验法分析(Bonnet & Vander Peer 2002).地理群体间的距离使用经纬度与坐标共同表示.最后,分子变异分析(analysis of molecular variance,AMOVA)用于估算地理隔离对病菌ITS单倍型分布的相对影响(Excoffier et al. 1992).用DnaSP 5.0软件(Librado & Rozas 2009)统计ITS序列的核苷酸多样性(nucleotide diversity,Pi)和群体间遗传分化指数(fixation index,FST);根据ITS序列的单倍型频率和核苷酸差异,用 TCS1.21软件(Clement et al. 2000)构建单倍型的网状亲缘关系图.采用DnaSP5.0软件对所有地理种群序列的单倍型错配进行分析.第二种类型的分析中,每个ITS单倍型序列都用于从GenBank中搜索近缘的序列.然后用Clustal W软件(Higgins et al. 1994)对我们的序列及搜索得到的序列进行排列,手工适当校正后用Mega 5.0软件进行分析.使用 Kimura双参数模型,排列中产生的空位(gap)被处理为缺失(missing)构建分子系统发育树(N-J tree),自展法(Bootstrap,1 000次重复)检验各分支的置信度(Tamura et al. 2004). ...
Colletotrichum species on grape in Guizhou and Yunnan provinces, China.
1
2013
... 油茶Camellia oleifera Abel.是原产于我国的食用木本油料树种.茶油中不饱和脂肪酸含量高,经常食用可降低血脂、肝脂、减轻血小板聚集力,防止血栓形成,对预防冠心病和防治动脉粥样硬化均有良好的作用(徐学兵 1995).近几年我国各油茶产区的炭疽病发生严重,引起严重落果、落蕾、枝梢枯死,甚至整株衰亡,造成重大经济损失(靳爱仙等 2009).目前,国内学者认为油茶炭疽病是由盘长孢状刺盘孢菌Colletotrichum gloeosporioides Penz.引起(周仲铭 1990;袁嗣令等 1997;杨光道等 2004;沈万芳 2008;刘伟 2012).我们通过分离病原菌、致病性测验、形态特征和多基因谱系分析,发现Colletotrichum fructicola Prihastuti, L. Cai & K.D. Hyde也能引起油茶炭疽病,而且是主要致病菌,其分布范围十分广阔.据Weir et al.(2012)报道,C. fructicola最早是在泰国的咖啡浆果上发现,随后在日本的沙梨、以色列的补血草、美国的葡萄和草莓以及德国的榕属植物上相继发现.在我国的贵州、云南等其他省份也发现该菌,其寄主包括辣椒、花生、灰莉、西红柿、冬青、葡萄和茶树等(杨友联2010;Peng et al. 2013;张艳敏 2012).目前,对C. fructicola的研究仅限于种名的确定.本研究拟分析C. fructicola种群遗传结构,这对控制油茶炭疽病具有重要意义.我们将用C. fructicola的ITS-5.8S区序列来分析以下5个方面的问题:第一,来自同一个地区的C. fructicola种群的个体间存在的变异;第二,ITS单倍型在不同地区中的分布规律;第三,C. fructicola不同的地理种群间是否具有显著的遗传分化?如果有,遗传分化的水平与地理距离是否存在明确的关系;第四,C. fructicola种群是否经历过种群扩张;第五,在系统分析中,当与相近种进行比较时,同一地理种群所有C. fructicola菌株是否会紧密地聚为一支. ...
Progress towards DNA barcoding of fungi.
1
2009
... 核糖体DNA内转录间隔区ITS(internal transcribed spacer)(包括5.8S)是介于18S和26S之间的非编码转录间隔区,承受的选择压力较小,进化速率快,能提供比较丰富的核苷酸变异位点和信息位点,且由于协同进化使得这个片段在不同的重复单元之间十分一致,此外ITS序列片段大小适中,易于测序和分析,近年来被广泛地应用于解决较低分类单元层次上的真菌系统发育问题(Seifert 2009;Bellemain et al. 2010).Sha et al.(2008)用ITS技术研究云南干巴菌Thelephora ganbajun的遗传多样性,利用通用引物ITS4和ITS5扩增该菌156个个体的ITS区域,结果表明不同地理来源的种群间变异为0.825,种群内变异为0.175,即该菌遗传分化较大.刘诗胤等(2012)利用ITS测序技术研究了浙江地区不同寄主来源炭疽菌的遗传多样性,结果表明不同菌株间的ITS具有丰富的多态性.刘芳等(2011)应用rDNA-ITS序列分析方法研究悬钩子皮下盘菌种内的遗传多样性,序列比较分析表明,悬钩子皮下盘菌ITS序列长度为443-456bp,G+C含量为48.7%-51.5%;不同菌株间的ITS区序列变异较大,该菌种内遗传多样性丰富.张振等(2009)对陕西省31 株木本植物炭疽菌的rDNA-ITS区序列进行了分析,发现炭疽菌rDNA的ITS区序列存在明显的差异,其ITS-rDNA序列多样性达0.1957,序列长度差异达94bp,通过聚类分析可将其分为不同的遗传类群.因此,炭疽菌rDNA的ITS区序列对种及种下专化型的划分具有一定的意义.本研究利用ITS序列,分析了我国果生刺盘孢菌的种群遗传结构,发现该病菌在我国油茶产区分布十分广泛,单倍型多样性较丰富,存在一定的遗传分化现象.Kelemu et al.(1997)对45个来自南美的圭亚那柱花草盘长孢状刺盘孢菌Colletotrichum gloeosporioides Penz.进行了RAPD遗传多态性分析,结果表明柱花草盘长孢状刺盘孢菌具有显著多态性,来自长期种植区的菌株的遗传多态性比新植区的丰富,这与本研究中,湖南天际岭和湖北随州这些超过50年树龄的地区病菌单倍型多样性丰富度一致. ...
Genetic diversity of the endemic gourmet mushroom Thelephora ganbajun from south western China.
1
2008
... 核糖体DNA内转录间隔区ITS(internal transcribed spacer)(包括5.8S)是介于18S和26S之间的非编码转录间隔区,承受的选择压力较小,进化速率快,能提供比较丰富的核苷酸变异位点和信息位点,且由于协同进化使得这个片段在不同的重复单元之间十分一致,此外ITS序列片段大小适中,易于测序和分析,近年来被广泛地应用于解决较低分类单元层次上的真菌系统发育问题(Seifert 2009;Bellemain et al. 2010).Sha et al.(2008)用ITS技术研究云南干巴菌Thelephora ganbajun的遗传多样性,利用通用引物ITS4和ITS5扩增该菌156个个体的ITS区域,结果表明不同地理来源的种群间变异为0.825,种群内变异为0.175,即该菌遗传分化较大.刘诗胤等(2012)利用ITS测序技术研究了浙江地区不同寄主来源炭疽菌的遗传多样性,结果表明不同菌株间的ITS具有丰富的多态性.刘芳等(2011)应用rDNA-ITS序列分析方法研究悬钩子皮下盘菌种内的遗传多样性,序列比较分析表明,悬钩子皮下盘菌ITS序列长度为443-456bp,G+C含量为48.7%-51.5%;不同菌株间的ITS区序列变异较大,该菌种内遗传多样性丰富.张振等(2009)对陕西省31 株木本植物炭疽菌的rDNA-ITS区序列进行了分析,发现炭疽菌rDNA的ITS区序列存在明显的差异,其ITS-rDNA序列多样性达0.1957,序列长度差异达94bp,通过聚类分析可将其分为不同的遗传类群.因此,炭疽菌rDNA的ITS区序列对种及种下专化型的划分具有一定的意义.本研究利用ITS序列,分析了我国果生刺盘孢菌的种群遗传结构,发现该病菌在我国油茶产区分布十分广泛,单倍型多样性较丰富,存在一定的遗传分化现象.Kelemu et al.(1997)对45个来自南美的圭亚那柱花草盘长孢状刺盘孢菌Colletotrichum gloeosporioides Penz.进行了RAPD遗传多态性分析,结果表明柱花草盘长孢状刺盘孢菌具有显著多态性,来自长期种植区的菌株的遗传多态性比新植区的丰富,这与本研究中,湖南天际岭和湖北随州这些超过50年树龄的地区病菌单倍型多样性丰富度一致. ...
Resistance mechanism of leave structure to and the influence of the stand and climatic factors to Colletotrichum gloeosporioides. Master Thesis,
0
2008
Prospects for inferring very large phylogenies by using the neighbor-joining method.
1
2004
... 采用BioEdit软件(Hall 1999)对DNA序列进行比对和编辑,同时辅以手工矫正.用Mega 5.0(Tamura et al. 2011)软件统计碱基组成、变异位点数等序列特征.使用GenALEx6(Peakall & Smouse 2006)分析各不同地区果生刺盘孢菌群体的ITS单倍型,研究:(1)各不同群体的单倍型多样性;(2)不同地区群体之间的遗传关系;(3)遗传分化与地理隔离的关系.其中,每个单独的群体及群体间的单倍型多样性和群体遗传分化FST值分开计算.遗传分化与地理距离的关系使用Mantel 检验法分析(Bonnet & Vander Peer 2002).地理群体间的距离使用经纬度与坐标共同表示.最后,分子变异分析(analysis of molecular variance,AMOVA)用于估算地理隔离对病菌ITS单倍型分布的相对影响(Excoffier et al. 1992).用DnaSP 5.0软件(Librado & Rozas 2009)统计ITS序列的核苷酸多样性(nucleotide diversity,Pi)和群体间遗传分化指数(fixation index,FST);根据ITS序列的单倍型频率和核苷酸差异,用 TCS1.21软件(Clement et al. 2000)构建单倍型的网状亲缘关系图.采用DnaSP5.0软件对所有地理种群序列的单倍型错配进行分析.第二种类型的分析中,每个ITS单倍型序列都用于从GenBank中搜索近缘的序列.然后用Clustal W软件(Higgins et al. 1994)对我们的序列及搜索得到的序列进行排列,手工适当校正后用Mega 5.0软件进行分析.使用 Kimura双参数模型,排列中产生的空位(gap)被处理为缺失(missing)构建分子系统发育树(N-J tree),自展法(Bootstrap,1 000次重复)检验各分支的置信度(Tamura et al. 2004). ...
MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods.
1
2011
... 采用BioEdit软件(Hall 1999)对DNA序列进行比对和编辑,同时辅以手工矫正.用Mega 5.0(Tamura et al. 2011)软件统计碱基组成、变异位点数等序列特征.使用GenALEx6(Peakall & Smouse 2006)分析各不同地区果生刺盘孢菌群体的ITS单倍型,研究:(1)各不同群体的单倍型多样性;(2)不同地区群体之间的遗传关系;(3)遗传分化与地理隔离的关系.其中,每个单独的群体及群体间的单倍型多样性和群体遗传分化FST值分开计算.遗传分化与地理距离的关系使用Mantel 检验法分析(Bonnet & Vander Peer 2002).地理群体间的距离使用经纬度与坐标共同表示.最后,分子变异分析(analysis of molecular variance,AMOVA)用于估算地理隔离对病菌ITS单倍型分布的相对影响(Excoffier et al. 1992).用DnaSP 5.0软件(Librado & Rozas 2009)统计ITS序列的核苷酸多样性(nucleotide diversity,Pi)和群体间遗传分化指数(fixation index,FST);根据ITS序列的单倍型频率和核苷酸差异,用 TCS1.21软件(Clement et al. 2000)构建单倍型的网状亲缘关系图.采用DnaSP5.0软件对所有地理种群序列的单倍型错配进行分析.第二种类型的分析中,每个ITS单倍型序列都用于从GenBank中搜索近缘的序列.然后用Clustal W软件(Higgins et al. 1994)对我们的序列及搜索得到的序列进行排列,手工适当校正后用Mega 5.0软件进行分析.使用 Kimura双参数模型,排列中产生的空位(gap)被处理为缺失(missing)构建分子系统发育树(N-J tree),自展法(Bootstrap,1 000次重复)检验各分支的置信度(Tamura et al. 2004). ...
The Colletotrichum gloeosporioides species complex.
1
2012
... 油茶Camellia oleifera Abel.是原产于我国的食用木本油料树种.茶油中不饱和脂肪酸含量高,经常食用可降低血脂、肝脂、减轻血小板聚集力,防止血栓形成,对预防冠心病和防治动脉粥样硬化均有良好的作用(徐学兵 1995).近几年我国各油茶产区的炭疽病发生严重,引起严重落果、落蕾、枝梢枯死,甚至整株衰亡,造成重大经济损失(靳爱仙等 2009).目前,国内学者认为油茶炭疽病是由盘长孢状刺盘孢菌Colletotrichum gloeosporioides Penz.引起(周仲铭 1990;袁嗣令等 1997;杨光道等 2004;沈万芳 2008;刘伟 2012).我们通过分离病原菌、致病性测验、形态特征和多基因谱系分析,发现Colletotrichum fructicola Prihastuti, L. Cai & K.D. Hyde也能引起油茶炭疽病,而且是主要致病菌,其分布范围十分广阔.据Weir et al.(2012)报道,C. fructicola最早是在泰国的咖啡浆果上发现,随后在日本的沙梨、以色列的补血草、美国的葡萄和草莓以及德国的榕属植物上相继发现.在我国的贵州、云南等其他省份也发现该菌,其寄主包括辣椒、花生、灰莉、西红柿、冬青、葡萄和茶树等(杨友联2010;Peng et al. 2013;张艳敏 2012).目前,对C. fructicola的研究仅限于种名的确定.本研究拟分析C. fructicola种群遗传结构,这对控制油茶炭疽病具有重要意义.我们将用C. fructicola的ITS-5.8S区序列来分析以下5个方面的问题:第一,来自同一个地区的C. fructicola种群的个体间存在的变异;第二,ITS单倍型在不同地区中的分布规律;第三,C. fructicola不同的地理种群间是否具有显著的遗传分化?如果有,遗传分化的水平与地理距离是否存在明确的关系;第四,C. fructicola种群是否经历过种群扩张;第五,在系统分析中,当与相近种进行比较时,同一地理种群所有C. fructicola菌株是否会紧密地聚为一支. ...





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}