Autophagy is a universal lysosomal degradation pathway in eukaryotic cells, and it plays an important role in maintaining normal cell functions by degrading excess or abnormal intracellular proteins, organelles, etc. During the past decade, the rapid progress of research in the field revealed that autophagy principally serves an adaptive role in diverse pathologies, including cancer, neurodegeneration, aging, and heart diseases. Moreover, autophagy also plays an essential role in filamentous fungi during growth, morphology, and development. Importantly, autophagy functions as a key factor during differentiation, such as pathogenic development, programmed cell death, and spore formation. This review focuses on nomenclature and detection of autophagy, and gives a glimpse on related studies about autophagy in fungi.
自噬是一种广泛存在于真核细胞内的依赖于溶酶体/液泡的降解途径,通过降解细胞内过多或异常的蛋白、细胞器等以维持正常的细胞功能。在自噬过程中,待降解的胞浆大分子物质或细胞器被双层膜泡结构包裹并通过细胞骨架微管网路系统运送至降解细胞器(溶酶体或液泡)中降解(Wang & Klionsky 2003),降解得到的产物分子如氨基酸等被运回细胞质内,重新利用。此外,自噬在某些情况下还能导致细胞死亡,因而被认为是区别于细胞凋亡的另一种细胞程序性死亡形式(Ⅱ型程序性死亡)(Edinger & Thompson 2004),细胞死亡命名委员会(Nomenclature Committee on Cell Death)分别在2009年和2011年讨论并推荐使用“自噬性细胞死亡”(autophagic cell death)这一概念来描述伴随着大量细胞质空泡化,往往(尽管不总是)表示自噬流(autophagic flux)增加的细胞死亡(Kroemer et al. 2009;Galluzzi et al. 2012)。“自噬性细胞死亡”这一概念暗示着自噬实际上可以执行细胞消亡,支持这一论证的证据之一是自噬似乎响应于体外选择的化疗药物而介导一些癌细胞的死亡(特别是当缺少重要的细胞凋亡调节因子如BAX和BAK或caspases时)(Grander et al. 2009)。同时,自噬作为细胞的管理程序,具有消除有缺陷的蛋白和细胞器,阻止异常蛋白的聚合积累和清除细胞内的病原体的功能,可能是防止衰老、癌症、神经退行性疾病和病原体感染的关键(Levine & Kroemer 2008)。已知自噬过程在几乎所有类型细胞的生理学和病理生理学方面都发挥着重要作用,自噬被认为是响应多种刺激的细胞反应(Codogno & Meijer 2005)。
2 自噬的分类
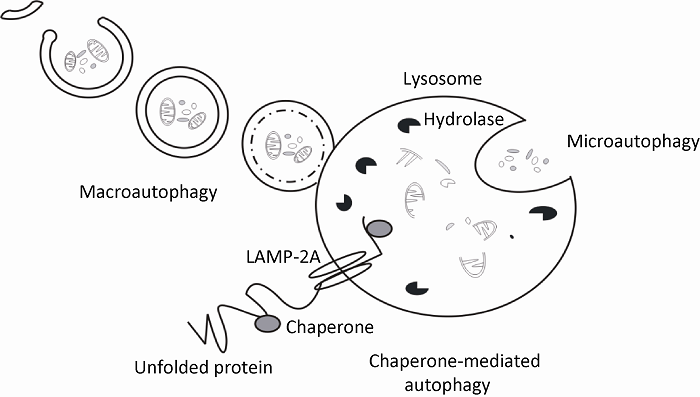
根据包裹底物分子及运送至降解细胞器方式的不同,可以将自噬分为巨自噬(macroautophagy),微自噬(microautophagy)和分子伴侣介导的自噬(chaperone-mediated autophagy)等3种(图1),其中巨自噬即通常所指的自噬,在本文此后的部分中,没有特殊标注的“自噬”均代表巨自噬(Mizushima et al. 2008)。
细胞自噬现象最初是由Christian de Duve等(Klionsky 2008)在20世纪60年代初发现的,当时并没有受到科学界的广泛关注,自噬研究的进展十分缓慢。对自噬现象较为透彻的研究首先在酿酒酵母Saccharomyces cerevisiae中进行,通过遗传学方法,到目前为止在酵母中已经发现了40多个与自噬相关的蛋白(表1),其中大部分在真核生物域内都十分保守,这说明自噬是一个进化保守的过程,其分子机制从酵母到哺乳动物十分相似。2003年,Klionsky et al.(2003)将编码这些蛋白的基因统一命名为Atg(AuTophaGy),用来代表自噬基因及其相对应的蛋白质。
Table 1
表1
表1 酵母中的Atg蛋白
Table 1 List of yeast Atg proteins
名称 Name
功能 Function
Atg1
丝/苏氨酸蛋白激酶,与Atg13和Atg17形成复合物,参与自噬的起始过程.
Protein serine/threonine kinase, forms a complex with Atg13 and Atg17, and participates in autophagy initiation.
Atg2
外周膜蛋白,参与自噬囊泡的形成,Atg9在自噬泡组装位点(PAS)及线粒体间的穿梭循环.
Peripheral membrane protein required for autophagic vesicle formation, involved in cycling of Atg9 from the PAS to the mitochondria.
Atg3
E2交联酶,参与Atg8与磷脂酰乙醇胺(PE)交联形成Atg8-PE的过程.
E2-like enzyme, plays a role in formation of Atg8-phosphatidylethanolamine conjugates.
Atg4
半胱氨酸蛋白酶,通过剪切碳末端的半胱氨酸残基或去除PE调节Atg8蛋白的脂化修饰.
Cysteine protease, cleaves the carboxy terminal arginine residue of Atg8, or PE from Atg8-PE conjugate.
这一系列的方法具有相对安全、简便,技术及仪器普及性高等优点,一般生化实验室即可达到应用的标准,因而被广泛地用来检测自噬。这一系列的检测方法包括:(1)电子显微镜(Eskelinen et al. 2011),利用电子显微镜分析自噬过程中多种膜结构的连续变化是一种即有效又必要的定性及定量分析方法;(2)Atg8/LC3的Western Blot分析(Mizushima & Yoshimori 2007),Atg8-PE/LC3-II是唯一一种可靠的与自噬体相关联的蛋白标记,通过对其含量的Western Blot分析来间接分析自噬体的含量;(3)荧光显微镜(Kabeya et al. 2000),将绿色荧光蛋白(GFP)或其他荧光蛋白的基因与Atg8/LC3蛋白的基因融合到一起,表达一个具有功能的融合蛋白,可以被应用于间接免疫荧光和荧光显微镜的方法检测自噬。
上述方法虽然能够直接或间接检测机体中自噬体的含量,但是我们不得不面对的一个问题是:自噬体的积累一定代表着自噬过程被诱导吗?实际上,自噬体消亡过程的减慢与自噬过程的诱导一样,也能引起自噬体的积累,而在这种情况下,底物分子不能正常的降解,反映的是一个自噬被抑制的过程。前者是通过抑制自噬体成熟成为自噬泡实现的,抑制是由于自噬体未能与胞内体或溶酶体融合,或者是融合后的低效降解而引起的。上面提到的检测自噬体的方法都具有一定程度的局限性,在具体应用时应谨慎。实际上,Klionsky et al.(2012)的千人签名的文章已经指出,必须检测自噬流。
5.2 通过自噬流的方法检测自噬
自噬过程不仅仅是自噬体的形成和消亡,更重要的是底物分子在整个自噬系统中的传递,即自噬流(autophagic flux)。通过检测底物分子在细胞中的传递来分析自噬过程更为恰当。通过自噬流检测自噬的方法包括:(1)自噬性蛋白质的降解,利用放射性氨基酸标记检测细胞内长寿命蛋白的降解来检测自噬过程(Pattingre et al. 2004);(2)LC3-II的周转周期(Tanida et al. 2005),使用蛋白酶抑制剂或改变溶酶体pH的巴弗洛霉素A1阻止溶酶体的降解,或使用阻断溶酶体与自噬体融合的试剂,使得LC3-II的含量不再变化,利用免疫印迹的方法推断LC3-II的周转周期来测量;(3)p62 western印迹(Mizushima & Yoshimori 2007),P62蛋白的作用是联系LC3和泛素化底物,p62被包含到完整的自噬体中,在自噬泡阶段被降解,最近的一项研究表明p62含量的增加与自噬的抑制相关,指出这种蛋白稳定状态下的含量反映了自噬过程的状态;(4)RFP-GFP联合荧光显微镜(Kimura et al. 2007)等。
自20世纪90年代初,Daniel J. Klionsky和Yoshinori Ohsumi等细胞生物学家以酿酒酵母为模式生物展开了自噬相关基因的筛选和细胞生物学的研究,经过二十多年的努力,酵母已经成为细胞自噬分子机制研究最为透彻的模式生物。细胞自噬既可以是非选择性的,也可以是选择性的。酵母中的选择性细胞自噬主要有cvt(cytoplasm to vacuole targeting)途径、线粒体自噬(mitophy)、过氧化物酶体自噬(pexophagy)等途径。经过多年的探索与积累,研究人员发明了多种多样的针对酵母细胞自噬的检测方法,如碱性磷酸酶法(Pho8△60测定法)。PHO8是酵母液泡中碱性磷酸酶的唯一编码基因,碱性磷酸酶是一种跨膜蛋白。其N端的60个氨基酸构成跨膜信号,负责前体pho8的一系列转运。为了利用Pho8来检测细胞自噬的水平,Noda et al.(1995)将其N端的60个氨基酸去掉,得到了Pho8△60蛋白。这样处理后获得的Pho8△60蛋白无法自主进入内质网,弥散分布在细胞质中,而当发生非选择性细胞自噬时,Pho8△60蛋白会被非选择性地包裹进入自噬体,从而运送到液泡并被激活,因而可以通过检测碱性磷酸酶的酶活性来检测细胞内自噬的发生水平。
6.2 丝状真菌与自噬
丝状真菌在人类社会中十分常见,在健康、食品、农业等方面都发挥着十分重要的作用,在自然条件及工业发酵条件下,丝状真菌持续经历着营养匮乏,在这种条件下,丝状真菌可能通过自噬来促进细胞存活及分化(Bartoszewska & Kiel 2011a)。值得注意的是,到目前为止,对于丝状真菌中选择性自噬途径的研究还不是十分透彻,仅有几篇关于过氧化物酶体自噬的报道(Amor et al. 2000;Asakura et al. 2009)。细胞自噬在丝状真菌的产孢、程序性细胞死亡、致病及促进细胞生存过程中发挥重要作用。在丝状真菌米曲霉Aspergillus oryzae中,Aoatg8、Aoatg13及Aoatg15的功能缺失将影响其产孢过程,说明自噬在该种真菌的无性生殖中发挥作用(Kikuma & Kitamoto 2011)。无独有偶,在其他丝状真菌中,自噬相关基因同样影响着产孢过程,如烟曲霉Aspergillus fumigatus中的Afatg1(Richie et al. 2007),以及稻瘟病菌Magnaporthe oryzae中的Moatg1、Moatg4、Moatg5、Moatg8和Moatg9(Liu et al. 2007)等。稻瘟病的致病过程起始于分生孢子的萌发,随后产生附着胞(appressoria)侵染宿主;附着胞的产生伴随着孢子结构的解体,这一过程需要通过自噬来回收孢子内的物质供给附着胞的生长(Veneault-Fourrey et al. 2006)。稻瘟病菌中Moatg1、Moatg2、Moatg3、Moatg4、Moatg5、Moatg6、Moatg7、Moatg8、Moatg9、Moatg10、Moatg12、Moatg13、Moatg15、Moatg16、Moatg17和Moatg18的缺失突变体不能引发稻瘟病,表明这些基因在稻瘟病菌致病过程中发挥作用;这种致病性的缺失是由于附着胞内甘油浓度的下降而导致细胞膨压的降低引起的(Lu et al. 2009)。而在大孢粪壳菌Sordaria macrospora中,Smatg8及Smatg4通过选择性及非选择性自噬参与调控营养生长、自噬体的发育及子囊袍子萌发等过程(Oliver & Stefanie 2013)。
许多丝状真菌对于植物来说是致病菌,农业上,这些真菌引起的真菌病害将导致农作物的减产及质量的下降。传统的防治真菌病害的药物存在着药物残留高,病菌易产生抗药性等问题,而现代农业逐渐强调低毒、低残留、高效及环境相容性。如果可以以真菌自噬为靶点,筛选出特异性抑制真菌自噬的化合物,从而抑制真菌对宿主的侵染,也许可以克服传统农药的弊端。另一方面,许多真菌次级代谢产物具有抗癌抗肿瘤作用。近年来的研究提供了越来越多的证据支持自噬是一种肿瘤抑制途径的观点(Levine 2007)。事实上,调节自噬和调节肿瘤发生的信号通路密切重叠,如TOR信号上游抑制子是抑癌基因,包括PTEN、TSC1和TSC2,刺激自噬过程,相反,激活致癌基因TOR的蛋白如Ⅰ类PI3K和Akt等则抑制自噬。尽管目前尚不清楚细胞存活/细胞死亡效应是否与自噬抑制肿瘤的作用有关,但是这些效应可能在癌症治疗过程中十分重要。针对ATG基因用siRNAs特异性的抑制自噬过程往往在这些实验中加速细胞死亡,而不是防止(Maiuri et al. 2007),说明自噬激活代表着一个细胞试图应对细胞毒性药物诱导产生的压力。这表明抑制自噬(而不是激活自噬)可能更有利于癌症治疗。C77,源于Gliocladium sp.的次生代谢产物,属于多硫代二酮哌嗪(epipolythiodioxopiperazines,ETPs)类化合物(Dong et al. 2005;Gardiner et al. 2005),具有广谱的生物活性。本实验室近期的研究结果表明,C77在一定程度上抑制乙肝病毒的复制,而这一过程可能是通过抑制乙肝细胞自噬过程引起的。丝状真菌中的自噬过程影响着次级代谢产物的生成及释放。产黄青霉Penicillium chrysogenum中atg1的缺失突变体在抑制了自噬过程的同时明显提高了青霉素的产量(Bartoszewska et al. 2011b)。因而,更好地了解自噬的分子机制将有利于我们以自噬为靶点寻求肿瘤治疗的突破点。
Novel system for monitoring autophagy in the yeast Saccharomyces cerevisiae.
Biochemical and Biophysical Research Communications, 210: 126-132
[27]
OliverV, StefanieP, 2013.
Autophagy genes Smatg8 and Smatg4 are required for fruiting-body development, vegetative growth and ascospore germination in the filamentous ascomycete Sordaria macrospora. Autophagy,
The catabolite inactivation of Aspergillus nidulans isocitrate lyase occurs by specific autophagy of peroxisomes.
1
2000
... 丝状真菌在人类社会中十分常见,在健康、食品、农业等方面都发挥着十分重要的作用,在自然条件及工业发酵条件下,丝状真菌持续经历着营养匮乏,在这种条件下,丝状真菌可能通过自噬来促进细胞存活及分化(Bartoszewska & Kiel 2011a).值得注意的是,到目前为止,对于丝状真菌中选择性自噬途径的研究还不是十分透彻,仅有几篇关于过氧化物酶体自噬的报道(Amor et al. 2000;Asakura et al. 2009).细胞自噬在丝状真菌的产孢、程序性细胞死亡、致病及促进细胞生存过程中发挥重要作用.在丝状真菌米曲霉Aspergillus oryzae中,Aoatg8、Aoatg13及Aoatg15的功能缺失将影响其产孢过程,说明自噬在该种真菌的无性生殖中发挥作用(Kikuma & Kitamoto 2011).无独有偶,在其他丝状真菌中,自噬相关基因同样影响着产孢过程,如烟曲霉Aspergillus fumigatus中的Afatg1(Richie et al. 2007),以及稻瘟病菌Magnaporthe oryzae中的Moatg1、Moatg4、Moatg5、Moatg8和Moatg9(Liu et al. 2007)等.稻瘟病的致病过程起始于分生孢子的萌发,随后产生附着胞(appressoria)侵染宿主;附着胞的产生伴随着孢子结构的解体,这一过程需要通过自噬来回收孢子内的物质供给附着胞的生长(Veneault-Fourrey et al. 2006).稻瘟病菌中Moatg1、Moatg2、Moatg3、Moatg4、Moatg5、Moatg6、Moatg7、Moatg8、Moatg9、Moatg10、Moatg12、Moatg13、Moatg15、Moatg16、Moatg17和Moatg18的缺失突变体不能引发稻瘟病,表明这些基因在稻瘟病菌致病过程中发挥作用;这种致病性的缺失是由于附着胞内甘油浓度的下降而导致细胞膨压的降低引起的(Lu et al. 2009).而在大孢粪壳菌Sordaria macrospora中,Smatg8及Smatg4通过选择性及非选择性自噬参与调控营养生长、自噬体的发育及子囊袍子萌发等过程(Oliver & Stefanie 2013). ...
Atg26-mediated pexophagy is required for host invasion by the plant pathogenic fungus Colletotrichum orbiculare.
1
2009
... 丝状真菌在人类社会中十分常见,在健康、食品、农业等方面都发挥着十分重要的作用,在自然条件及工业发酵条件下,丝状真菌持续经历着营养匮乏,在这种条件下,丝状真菌可能通过自噬来促进细胞存活及分化(Bartoszewska & Kiel 2011a).值得注意的是,到目前为止,对于丝状真菌中选择性自噬途径的研究还不是十分透彻,仅有几篇关于过氧化物酶体自噬的报道(Amor et al. 2000;Asakura et al. 2009).细胞自噬在丝状真菌的产孢、程序性细胞死亡、致病及促进细胞生存过程中发挥重要作用.在丝状真菌米曲霉Aspergillus oryzae中,Aoatg8、Aoatg13及Aoatg15的功能缺失将影响其产孢过程,说明自噬在该种真菌的无性生殖中发挥作用(Kikuma & Kitamoto 2011).无独有偶,在其他丝状真菌中,自噬相关基因同样影响着产孢过程,如烟曲霉Aspergillus fumigatus中的Afatg1(Richie et al. 2007),以及稻瘟病菌Magnaporthe oryzae中的Moatg1、Moatg4、Moatg5、Moatg8和Moatg9(Liu et al. 2007)等.稻瘟病的致病过程起始于分生孢子的萌发,随后产生附着胞(appressoria)侵染宿主;附着胞的产生伴随着孢子结构的解体,这一过程需要通过自噬来回收孢子内的物质供给附着胞的生长(Veneault-Fourrey et al. 2006).稻瘟病菌中Moatg1、Moatg2、Moatg3、Moatg4、Moatg5、Moatg6、Moatg7、Moatg8、Moatg9、Moatg10、Moatg12、Moatg13、Moatg15、Moatg16、Moatg17和Moatg18的缺失突变体不能引发稻瘟病,表明这些基因在稻瘟病菌致病过程中发挥作用;这种致病性的缺失是由于附着胞内甘油浓度的下降而导致细胞膨压的降低引起的(Lu et al. 2009).而在大孢粪壳菌Sordaria macrospora中,Smatg8及Smatg4通过选择性及非选择性自噬参与调控营养生长、自噬体的发育及子囊袍子萌发等过程(Oliver & Stefanie 2013). ...
a. The role of macroautophagy in development of filamentous fungi
1
2011
... 丝状真菌在人类社会中十分常见,在健康、食品、农业等方面都发挥着十分重要的作用,在自然条件及工业发酵条件下,丝状真菌持续经历着营养匮乏,在这种条件下,丝状真菌可能通过自噬来促进细胞存活及分化(Bartoszewska & Kiel 2011a).值得注意的是,到目前为止,对于丝状真菌中选择性自噬途径的研究还不是十分透彻,仅有几篇关于过氧化物酶体自噬的报道(Amor et al. 2000;Asakura et al. 2009).细胞自噬在丝状真菌的产孢、程序性细胞死亡、致病及促进细胞生存过程中发挥重要作用.在丝状真菌米曲霉Aspergillus oryzae中,Aoatg8、Aoatg13及Aoatg15的功能缺失将影响其产孢过程,说明自噬在该种真菌的无性生殖中发挥作用(Kikuma & Kitamoto 2011).无独有偶,在其他丝状真菌中,自噬相关基因同样影响着产孢过程,如烟曲霉Aspergillus fumigatus中的Afatg1(Richie et al. 2007),以及稻瘟病菌Magnaporthe oryzae中的Moatg1、Moatg4、Moatg5、Moatg8和Moatg9(Liu et al. 2007)等.稻瘟病的致病过程起始于分生孢子的萌发,随后产生附着胞(appressoria)侵染宿主;附着胞的产生伴随着孢子结构的解体,这一过程需要通过自噬来回收孢子内的物质供给附着胞的生长(Veneault-Fourrey et al. 2006).稻瘟病菌中Moatg1、Moatg2、Moatg3、Moatg4、Moatg5、Moatg6、Moatg7、Moatg8、Moatg9、Moatg10、Moatg12、Moatg13、Moatg15、Moatg16、Moatg17和Moatg18的缺失突变体不能引发稻瘟病,表明这些基因在稻瘟病菌致病过程中发挥作用;这种致病性的缺失是由于附着胞内甘油浓度的下降而导致细胞膨压的降低引起的(Lu et al. 2009).而在大孢粪壳菌Sordaria macrospora中,Smatg8及Smatg4通过选择性及非选择性自噬参与调控营养生长、自噬体的发育及子囊袍子萌发等过程(Oliver & Stefanie 2013). ...
b. Autophagy deficiency promotes β-lactam production in Penicillium chrysogenum.
1
2011
... 许多丝状真菌对于植物来说是致病菌,农业上,这些真菌引起的真菌病害将导致农作物的减产及质量的下降.传统的防治真菌病害的药物存在着药物残留高,病菌易产生抗药性等问题,而现代农业逐渐强调低毒、低残留、高效及环境相容性.如果可以以真菌自噬为靶点,筛选出特异性抑制真菌自噬的化合物,从而抑制真菌对宿主的侵染,也许可以克服传统农药的弊端.另一方面,许多真菌次级代谢产物具有抗癌抗肿瘤作用.近年来的研究提供了越来越多的证据支持自噬是一种肿瘤抑制途径的观点(Levine 2007).事实上,调节自噬和调节肿瘤发生的信号通路密切重叠,如TOR信号上游抑制子是抑癌基因,包括PTEN、TSC1和TSC2,刺激自噬过程,相反,激活致癌基因TOR的蛋白如Ⅰ类PI3K和Akt等则抑制自噬.尽管目前尚不清楚细胞存活/细胞死亡效应是否与自噬抑制肿瘤的作用有关,但是这些效应可能在癌症治疗过程中十分重要.针对ATG基因用siRNAs特异性的抑制自噬过程往往在这些实验中加速细胞死亡,而不是防止(Maiuri et al. 2007),说明自噬激活代表着一个细胞试图应对细胞毒性药物诱导产生的压力.这表明抑制自噬(而不是激活自噬)可能更有利于癌症治疗.C77,源于Gliocladium sp.的次生代谢产物,属于多硫代二酮哌嗪(epipolythiodioxopiperazines,ETPs)类化合物(Dong et al. 2005;Gardiner et al. 2005),具有广谱的生物活性.本实验室近期的研究结果表明,C77在一定程度上抑制乙肝病毒的复制,而这一过程可能是通过抑制乙肝细胞自噬过程引起的.丝状真菌中的自噬过程影响着次级代谢产物的生成及释放.产黄青霉Penicillium chrysogenum中atg1的缺失突变体在抑制了自噬过程的同时明显提高了青霉素的产量(Bartoszewska et al. 2011b).因而,更好地了解自噬的分子机制将有利于我们以自噬为靶点寻求肿瘤治疗的突破点. ...
Autophagy and signaling: their role in cell survival and cell death.
1
2005
... 自噬是一种广泛存在于真核细胞内的依赖于溶酶体/液泡的降解途径,通过降解细胞内过多或异常的蛋白、细胞器等以维持正常的细胞功能.在自噬过程中,待降解的胞浆大分子物质或细胞器被双层膜泡结构包裹并通过细胞骨架微管网路系统运送至降解细胞器(溶酶体或液泡)中降解(Wang & Klionsky 2003),降解得到的产物分子如氨基酸等被运回细胞质内,重新利用.此外,自噬在某些情况下还能导致细胞死亡,因而被认为是区别于细胞凋亡的另一种细胞程序性死亡形式(Ⅱ型程序性死亡)(Edinger & Thompson 2004),细胞死亡命名委员会(Nomenclature Committee on Cell Death)分别在2009年和2011年讨论并推荐使用“自噬性细胞死亡”(autophagic cell death)这一概念来描述伴随着大量细胞质空泡化,往往(尽管不总是)表示自噬流(autophagic flux)增加的细胞死亡(Kroemer et al. 2009;Galluzzi et al. 2012).“自噬性细胞死亡”这一概念暗示着自噬实际上可以执行细胞消亡,支持这一论证的证据之一是自噬似乎响应于体外选择的化疗药物而介导一些癌细胞的死亡(特别是当缺少重要的细胞凋亡调节因子如BAX和BAK或caspases时)(Grander et al. 2009).同时,自噬作为细胞的管理程序,具有消除有缺陷的蛋白和细胞器,阻止异常蛋白的聚合积累和清除细胞内的病原体的功能,可能是防止衰老、癌症、神经退行性疾病和病原体感染的关键(Levine & Kroemer 2008).已知自噬过程在几乎所有类型细胞的生理学和病理生理学方面都发挥着重要作用,自噬被认为是响应多种刺激的细胞反应(Codogno & Meijer 2005). ...
Nematicidal epipolysulfanyldioxopiperazines from Gliocladium roseum.
1
2005
... 许多丝状真菌对于植物来说是致病菌,农业上,这些真菌引起的真菌病害将导致农作物的减产及质量的下降.传统的防治真菌病害的药物存在着药物残留高,病菌易产生抗药性等问题,而现代农业逐渐强调低毒、低残留、高效及环境相容性.如果可以以真菌自噬为靶点,筛选出特异性抑制真菌自噬的化合物,从而抑制真菌对宿主的侵染,也许可以克服传统农药的弊端.另一方面,许多真菌次级代谢产物具有抗癌抗肿瘤作用.近年来的研究提供了越来越多的证据支持自噬是一种肿瘤抑制途径的观点(Levine 2007).事实上,调节自噬和调节肿瘤发生的信号通路密切重叠,如TOR信号上游抑制子是抑癌基因,包括PTEN、TSC1和TSC2,刺激自噬过程,相反,激活致癌基因TOR的蛋白如Ⅰ类PI3K和Akt等则抑制自噬.尽管目前尚不清楚细胞存活/细胞死亡效应是否与自噬抑制肿瘤的作用有关,但是这些效应可能在癌症治疗过程中十分重要.针对ATG基因用siRNAs特异性的抑制自噬过程往往在这些实验中加速细胞死亡,而不是防止(Maiuri et al. 2007),说明自噬激活代表着一个细胞试图应对细胞毒性药物诱导产生的压力.这表明抑制自噬(而不是激活自噬)可能更有利于癌症治疗.C77,源于Gliocladium sp.的次生代谢产物,属于多硫代二酮哌嗪(epipolythiodioxopiperazines,ETPs)类化合物(Dong et al. 2005;Gardiner et al. 2005),具有广谱的生物活性.本实验室近期的研究结果表明,C77在一定程度上抑制乙肝病毒的复制,而这一过程可能是通过抑制乙肝细胞自噬过程引起的.丝状真菌中的自噬过程影响着次级代谢产物的生成及释放.产黄青霉Penicillium chrysogenum中atg1的缺失突变体在抑制了自噬过程的同时明显提高了青霉素的产量(Bartoszewska et al. 2011b).因而,更好地了解自噬的分子机制将有利于我们以自噬为靶点寻求肿瘤治疗的突破点. ...
Death by design: apoptosis, necrosis and autophagy.
1
2004
... 自噬是一种广泛存在于真核细胞内的依赖于溶酶体/液泡的降解途径,通过降解细胞内过多或异常的蛋白、细胞器等以维持正常的细胞功能.在自噬过程中,待降解的胞浆大分子物质或细胞器被双层膜泡结构包裹并通过细胞骨架微管网路系统运送至降解细胞器(溶酶体或液泡)中降解(Wang & Klionsky 2003),降解得到的产物分子如氨基酸等被运回细胞质内,重新利用.此外,自噬在某些情况下还能导致细胞死亡,因而被认为是区别于细胞凋亡的另一种细胞程序性死亡形式(Ⅱ型程序性死亡)(Edinger & Thompson 2004),细胞死亡命名委员会(Nomenclature Committee on Cell Death)分别在2009年和2011年讨论并推荐使用“自噬性细胞死亡”(autophagic cell death)这一概念来描述伴随着大量细胞质空泡化,往往(尽管不总是)表示自噬流(autophagic flux)增加的细胞死亡(Kroemer et al. 2009;Galluzzi et al. 2012).“自噬性细胞死亡”这一概念暗示着自噬实际上可以执行细胞消亡,支持这一论证的证据之一是自噬似乎响应于体外选择的化疗药物而介导一些癌细胞的死亡(特别是当缺少重要的细胞凋亡调节因子如BAX和BAK或caspases时)(Grander et al. 2009).同时,自噬作为细胞的管理程序,具有消除有缺陷的蛋白和细胞器,阻止异常蛋白的聚合积累和清除细胞内的病原体的功能,可能是防止衰老、癌症、神经退行性疾病和病原体感染的关键(Levine & Kroemer 2008).已知自噬过程在几乎所有类型细胞的生理学和病理生理学方面都发挥着重要作用,自噬被认为是响应多种刺激的细胞反应(Codogno & Meijer 2005). ...
Seeing is believing: the impact of electron microscopy on autophagy research.
1
2011
... 这一系列的方法具有相对安全、简便,技术及仪器普及性高等优点,一般生化实验室即可达到应用的标准,因而被广泛地用来检测自噬.这一系列的检测方法包括:(1)电子显微镜(Eskelinen et al. 2011),利用电子显微镜分析自噬过程中多种膜结构的连续变化是一种即有效又必要的定性及定量分析方法;(2)Atg8/LC3的Western Blot分析(Mizushima & Yoshimori 2007),Atg8-PE/LC3-II是唯一一种可靠的与自噬体相关联的蛋白标记,通过对其含量的Western Blot分析来间接分析自噬体的含量;(3)荧光显微镜(Kabeya et al. 2000),将绿色荧光蛋白(GFP)或其他荧光蛋白的基因与Atg8/LC3蛋白的基因融合到一起,表达一个具有功能的融合蛋白,可以被应用于间接免疫荧光和荧光显微镜的方法检测自噬. ...
Molecular definitions of cell death subroutines: recommendations of the Nomenclature Committee on Cell Death 2012.
1
2012
... 自噬是一种广泛存在于真核细胞内的依赖于溶酶体/液泡的降解途径,通过降解细胞内过多或异常的蛋白、细胞器等以维持正常的细胞功能.在自噬过程中,待降解的胞浆大分子物质或细胞器被双层膜泡结构包裹并通过细胞骨架微管网路系统运送至降解细胞器(溶酶体或液泡)中降解(Wang & Klionsky 2003),降解得到的产物分子如氨基酸等被运回细胞质内,重新利用.此外,自噬在某些情况下还能导致细胞死亡,因而被认为是区别于细胞凋亡的另一种细胞程序性死亡形式(Ⅱ型程序性死亡)(Edinger & Thompson 2004),细胞死亡命名委员会(Nomenclature Committee on Cell Death)分别在2009年和2011年讨论并推荐使用“自噬性细胞死亡”(autophagic cell death)这一概念来描述伴随着大量细胞质空泡化,往往(尽管不总是)表示自噬流(autophagic flux)增加的细胞死亡(Kroemer et al. 2009;Galluzzi et al. 2012).“自噬性细胞死亡”这一概念暗示着自噬实际上可以执行细胞消亡,支持这一论证的证据之一是自噬似乎响应于体外选择的化疗药物而介导一些癌细胞的死亡(特别是当缺少重要的细胞凋亡调节因子如BAX和BAK或caspases时)(Grander et al. 2009).同时,自噬作为细胞的管理程序,具有消除有缺陷的蛋白和细胞器,阻止异常蛋白的聚合积累和清除细胞内的病原体的功能,可能是防止衰老、癌症、神经退行性疾病和病原体感染的关键(Levine & Kroemer 2008).已知自噬过程在几乎所有类型细胞的生理学和病理生理学方面都发挥着重要作用,自噬被认为是响应多种刺激的细胞反应(Codogno & Meijer 2005). ...
The epipolythiodioxopiperazine (ETP) class of fungal toxins: distribution, mode of action, functions and biosynthesis.
1
2005
... 许多丝状真菌对于植物来说是致病菌,农业上,这些真菌引起的真菌病害将导致农作物的减产及质量的下降.传统的防治真菌病害的药物存在着药物残留高,病菌易产生抗药性等问题,而现代农业逐渐强调低毒、低残留、高效及环境相容性.如果可以以真菌自噬为靶点,筛选出特异性抑制真菌自噬的化合物,从而抑制真菌对宿主的侵染,也许可以克服传统农药的弊端.另一方面,许多真菌次级代谢产物具有抗癌抗肿瘤作用.近年来的研究提供了越来越多的证据支持自噬是一种肿瘤抑制途径的观点(Levine 2007).事实上,调节自噬和调节肿瘤发生的信号通路密切重叠,如TOR信号上游抑制子是抑癌基因,包括PTEN、TSC1和TSC2,刺激自噬过程,相反,激活致癌基因TOR的蛋白如Ⅰ类PI3K和Akt等则抑制自噬.尽管目前尚不清楚细胞存活/细胞死亡效应是否与自噬抑制肿瘤的作用有关,但是这些效应可能在癌症治疗过程中十分重要.针对ATG基因用siRNAs特异性的抑制自噬过程往往在这些实验中加速细胞死亡,而不是防止(Maiuri et al. 2007),说明自噬激活代表着一个细胞试图应对细胞毒性药物诱导产生的压力.这表明抑制自噬(而不是激活自噬)可能更有利于癌症治疗.C77,源于Gliocladium sp.的次生代谢产物,属于多硫代二酮哌嗪(epipolythiodioxopiperazines,ETPs)类化合物(Dong et al. 2005;Gardiner et al. 2005),具有广谱的生物活性.本实验室近期的研究结果表明,C77在一定程度上抑制乙肝病毒的复制,而这一过程可能是通过抑制乙肝细胞自噬过程引起的.丝状真菌中的自噬过程影响着次级代谢产物的生成及释放.产黄青霉Penicillium chrysogenum中atg1的缺失突变体在抑制了自噬过程的同时明显提高了青霉素的产量(Bartoszewska et al. 2011b).因而,更好地了解自噬的分子机制将有利于我们以自噬为靶点寻求肿瘤治疗的突破点. ...
Autophagy as the main means of cytotoxicity by glucocorticoids in hematological malignancies.
1
2009
... 自噬是一种广泛存在于真核细胞内的依赖于溶酶体/液泡的降解途径,通过降解细胞内过多或异常的蛋白、细胞器等以维持正常的细胞功能.在自噬过程中,待降解的胞浆大分子物质或细胞器被双层膜泡结构包裹并通过细胞骨架微管网路系统运送至降解细胞器(溶酶体或液泡)中降解(Wang & Klionsky 2003),降解得到的产物分子如氨基酸等被运回细胞质内,重新利用.此外,自噬在某些情况下还能导致细胞死亡,因而被认为是区别于细胞凋亡的另一种细胞程序性死亡形式(Ⅱ型程序性死亡)(Edinger & Thompson 2004),细胞死亡命名委员会(Nomenclature Committee on Cell Death)分别在2009年和2011年讨论并推荐使用“自噬性细胞死亡”(autophagic cell death)这一概念来描述伴随着大量细胞质空泡化,往往(尽管不总是)表示自噬流(autophagic flux)增加的细胞死亡(Kroemer et al. 2009;Galluzzi et al. 2012).“自噬性细胞死亡”这一概念暗示着自噬实际上可以执行细胞消亡,支持这一论证的证据之一是自噬似乎响应于体外选择的化疗药物而介导一些癌细胞的死亡(特别是当缺少重要的细胞凋亡调节因子如BAX和BAK或caspases时)(Grander et al. 2009).同时,自噬作为细胞的管理程序,具有消除有缺陷的蛋白和细胞器,阻止异常蛋白的聚合积累和清除细胞内的病原体的功能,可能是防止衰老、癌症、神经退行性疾病和病原体感染的关键(Levine & Kroemer 2008).已知自噬过程在几乎所有类型细胞的生理学和病理生理学方面都发挥着重要作用,自噬被认为是响应多种刺激的细胞反应(Codogno & Meijer 2005). ...
LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing.
1
2000
... 这一系列的方法具有相对安全、简便,技术及仪器普及性高等优点,一般生化实验室即可达到应用的标准,因而被广泛地用来检测自噬.这一系列的检测方法包括:(1)电子显微镜(Eskelinen et al. 2011),利用电子显微镜分析自噬过程中多种膜结构的连续变化是一种即有效又必要的定性及定量分析方法;(2)Atg8/LC3的Western Blot分析(Mizushima & Yoshimori 2007),Atg8-PE/LC3-II是唯一一种可靠的与自噬体相关联的蛋白标记,通过对其含量的Western Blot分析来间接分析自噬体的含量;(3)荧光显微镜(Kabeya et al. 2000),将绿色荧光蛋白(GFP)或其他荧光蛋白的基因与Atg8/LC3蛋白的基因融合到一起,表达一个具有功能的融合蛋白,可以被应用于间接免疫荧光和荧光显微镜的方法检测自噬. ...
Analysis of autophagy in Aspergillus oryzae by disruption of Aoatg13, Aoatg4, and Aoatg15 genes.
1
2011
... 丝状真菌在人类社会中十分常见,在健康、食品、农业等方面都发挥着十分重要的作用,在自然条件及工业发酵条件下,丝状真菌持续经历着营养匮乏,在这种条件下,丝状真菌可能通过自噬来促进细胞存活及分化(Bartoszewska & Kiel 2011a).值得注意的是,到目前为止,对于丝状真菌中选择性自噬途径的研究还不是十分透彻,仅有几篇关于过氧化物酶体自噬的报道(Amor et al. 2000;Asakura et al. 2009).细胞自噬在丝状真菌的产孢、程序性细胞死亡、致病及促进细胞生存过程中发挥重要作用.在丝状真菌米曲霉Aspergillus oryzae中,Aoatg8、Aoatg13及Aoatg15的功能缺失将影响其产孢过程,说明自噬在该种真菌的无性生殖中发挥作用(Kikuma & Kitamoto 2011).无独有偶,在其他丝状真菌中,自噬相关基因同样影响着产孢过程,如烟曲霉Aspergillus fumigatus中的Afatg1(Richie et al. 2007),以及稻瘟病菌Magnaporthe oryzae中的Moatg1、Moatg4、Moatg5、Moatg8和Moatg9(Liu et al. 2007)等.稻瘟病的致病过程起始于分生孢子的萌发,随后产生附着胞(appressoria)侵染宿主;附着胞的产生伴随着孢子结构的解体,这一过程需要通过自噬来回收孢子内的物质供给附着胞的生长(Veneault-Fourrey et al. 2006).稻瘟病菌中Moatg1、Moatg2、Moatg3、Moatg4、Moatg5、Moatg6、Moatg7、Moatg8、Moatg9、Moatg10、Moatg12、Moatg13、Moatg15、Moatg16、Moatg17和Moatg18的缺失突变体不能引发稻瘟病,表明这些基因在稻瘟病菌致病过程中发挥作用;这种致病性的缺失是由于附着胞内甘油浓度的下降而导致细胞膨压的降低引起的(Lu et al. 2009).而在大孢粪壳菌Sordaria macrospora中,Smatg8及Smatg4通过选择性及非选择性自噬参与调控营养生长、自噬体的发育及子囊袍子萌发等过程(Oliver & Stefanie 2013). ...
Dissection of the autophagosome maturation process by a novel reporter protein, tandem fluorescent-tagged LC3.
1
2007
... 自噬过程不仅仅是自噬体的形成和消亡,更重要的是底物分子在整个自噬系统中的传递,即自噬流(autophagic flux).通过检测底物分子在细胞中的传递来分析自噬过程更为恰当.通过自噬流检测自噬的方法包括:(1)自噬性蛋白质的降解,利用放射性氨基酸标记检测细胞内长寿命蛋白的降解来检测自噬过程(Pattingre et al. 2004);(2)LC3-II的周转周期(Tanida et al. 2005),使用蛋白酶抑制剂或改变溶酶体pH的巴弗洛霉素A1阻止溶酶体的降解,或使用阻断溶酶体与自噬体融合的试剂,使得LC3-II的含量不再变化,利用免疫印迹的方法推断LC3-II的周转周期来测量;(3)p62 western印迹(Mizushima & Yoshimori 2007),P62蛋白的作用是联系LC3和泛素化底物,p62被包含到完整的自噬体中,在自噬泡阶段被降解,最近的一项研究表明p62含量的增加与自噬的抑制相关,指出这种蛋白稳定状态下的含量反映了自噬过程的状态;(4)RFP-GFP联合荧光显微镜(Kimura et al. 2007)等. ...
Autophagy revisited: a conversation with Christian de Duve.
1
2008
... 细胞自噬现象最初是由Christian de Duve等(Klionsky 2008)在20世纪60年代初发现的,当时并没有受到科学界的广泛关注,自噬研究的进展十分缓慢.对自噬现象较为透彻的研究首先在酿酒酵母Saccharomyces cerevisiae中进行,通过遗传学方法,到目前为止在酵母中已经发现了40多个与自噬相关的蛋白(表1),其中大部分在真核生物域内都十分保守,这说明自噬是一个进化保守的过程,其分子机制从酵母到哺乳动物十分相似.2003年,Klionsky et al.(2003)将编码这些蛋白的基因统一命名为Atg(AuTophaGy),用来代表自噬基因及其相对应的蛋白质. ...
Guidelines for the use and interpretation of assays for monitoring autophagy.
0
2012
A unified nomenclature for yeast autophagy-related genes.
0
2003
Classification of cell death: recommendations of the Nomenclature Committee on Cell Death 2009.
1
2009
... 自噬是一种广泛存在于真核细胞内的依赖于溶酶体/液泡的降解途径,通过降解细胞内过多或异常的蛋白、细胞器等以维持正常的细胞功能.在自噬过程中,待降解的胞浆大分子物质或细胞器被双层膜泡结构包裹并通过细胞骨架微管网路系统运送至降解细胞器(溶酶体或液泡)中降解(Wang & Klionsky 2003),降解得到的产物分子如氨基酸等被运回细胞质内,重新利用.此外,自噬在某些情况下还能导致细胞死亡,因而被认为是区别于细胞凋亡的另一种细胞程序性死亡形式(Ⅱ型程序性死亡)(Edinger & Thompson 2004),细胞死亡命名委员会(Nomenclature Committee on Cell Death)分别在2009年和2011年讨论并推荐使用“自噬性细胞死亡”(autophagic cell death)这一概念来描述伴随着大量细胞质空泡化,往往(尽管不总是)表示自噬流(autophagic flux)增加的细胞死亡(Kroemer et al. 2009;Galluzzi et al. 2012).“自噬性细胞死亡”这一概念暗示着自噬实际上可以执行细胞消亡,支持这一论证的证据之一是自噬似乎响应于体外选择的化疗药物而介导一些癌细胞的死亡(特别是当缺少重要的细胞凋亡调节因子如BAX和BAK或caspases时)(Grander et al. 2009).同时,自噬作为细胞的管理程序,具有消除有缺陷的蛋白和细胞器,阻止异常蛋白的聚合积累和清除细胞内的病原体的功能,可能是防止衰老、癌症、神经退行性疾病和病原体感染的关键(Levine & Kroemer 2008).已知自噬过程在几乎所有类型细胞的生理学和病理生理学方面都发挥着重要作用,自噬被认为是响应多种刺激的细胞反应(Codogno & Meijer 2005). ...
Cell biology: autophagy and cancer.
1
2007
... 许多丝状真菌对于植物来说是致病菌,农业上,这些真菌引起的真菌病害将导致农作物的减产及质量的下降.传统的防治真菌病害的药物存在着药物残留高,病菌易产生抗药性等问题,而现代农业逐渐强调低毒、低残留、高效及环境相容性.如果可以以真菌自噬为靶点,筛选出特异性抑制真菌自噬的化合物,从而抑制真菌对宿主的侵染,也许可以克服传统农药的弊端.另一方面,许多真菌次级代谢产物具有抗癌抗肿瘤作用.近年来的研究提供了越来越多的证据支持自噬是一种肿瘤抑制途径的观点(Levine 2007).事实上,调节自噬和调节肿瘤发生的信号通路密切重叠,如TOR信号上游抑制子是抑癌基因,包括PTEN、TSC1和TSC2,刺激自噬过程,相反,激活致癌基因TOR的蛋白如Ⅰ类PI3K和Akt等则抑制自噬.尽管目前尚不清楚细胞存活/细胞死亡效应是否与自噬抑制肿瘤的作用有关,但是这些效应可能在癌症治疗过程中十分重要.针对ATG基因用siRNAs特异性的抑制自噬过程往往在这些实验中加速细胞死亡,而不是防止(Maiuri et al. 2007),说明自噬激活代表着一个细胞试图应对细胞毒性药物诱导产生的压力.这表明抑制自噬(而不是激活自噬)可能更有利于癌症治疗.C77,源于Gliocladium sp.的次生代谢产物,属于多硫代二酮哌嗪(epipolythiodioxopiperazines,ETPs)类化合物(Dong et al. 2005;Gardiner et al. 2005),具有广谱的生物活性.本实验室近期的研究结果表明,C77在一定程度上抑制乙肝病毒的复制,而这一过程可能是通过抑制乙肝细胞自噬过程引起的.丝状真菌中的自噬过程影响着次级代谢产物的生成及释放.产黄青霉Penicillium chrysogenum中atg1的缺失突变体在抑制了自噬过程的同时明显提高了青霉素的产量(Bartoszewska et al. 2011b).因而,更好地了解自噬的分子机制将有利于我们以自噬为靶点寻求肿瘤治疗的突破点. ...
Autophagy in the pathogenesis of disease.
1
2008
... 自噬是一种广泛存在于真核细胞内的依赖于溶酶体/液泡的降解途径,通过降解细胞内过多或异常的蛋白、细胞器等以维持正常的细胞功能.在自噬过程中,待降解的胞浆大分子物质或细胞器被双层膜泡结构包裹并通过细胞骨架微管网路系统运送至降解细胞器(溶酶体或液泡)中降解(Wang & Klionsky 2003),降解得到的产物分子如氨基酸等被运回细胞质内,重新利用.此外,自噬在某些情况下还能导致细胞死亡,因而被认为是区别于细胞凋亡的另一种细胞程序性死亡形式(Ⅱ型程序性死亡)(Edinger & Thompson 2004),细胞死亡命名委员会(Nomenclature Committee on Cell Death)分别在2009年和2011年讨论并推荐使用“自噬性细胞死亡”(autophagic cell death)这一概念来描述伴随着大量细胞质空泡化,往往(尽管不总是)表示自噬流(autophagic flux)增加的细胞死亡(Kroemer et al. 2009;Galluzzi et al. 2012).“自噬性细胞死亡”这一概念暗示着自噬实际上可以执行细胞消亡,支持这一论证的证据之一是自噬似乎响应于体外选择的化疗药物而介导一些癌细胞的死亡(特别是当缺少重要的细胞凋亡调节因子如BAX和BAK或caspases时)(Grander et al. 2009).同时,自噬作为细胞的管理程序,具有消除有缺陷的蛋白和细胞器,阻止异常蛋白的聚合积累和清除细胞内的病原体的功能,可能是防止衰老、癌症、神经退行性疾病和病原体感染的关键(Levine & Kroemer 2008).已知自噬过程在几乎所有类型细胞的生理学和病理生理学方面都发挥着重要作用,自噬被认为是响应多种刺激的细胞反应(Codogno & Meijer 2005). ...
Involvement of a Magnaporthe grisea serine/threonine kinase gene, MgATG1, in appressorium turgor and pathogenesis.
0
2007
An autophagy gene, MgATG5, is required for cell differentiation and pathogenesis in Magnaporthe oryzae.
1
2009
... 丝状真菌在人类社会中十分常见,在健康、食品、农业等方面都发挥着十分重要的作用,在自然条件及工业发酵条件下,丝状真菌持续经历着营养匮乏,在这种条件下,丝状真菌可能通过自噬来促进细胞存活及分化(Bartoszewska & Kiel 2011a).值得注意的是,到目前为止,对于丝状真菌中选择性自噬途径的研究还不是十分透彻,仅有几篇关于过氧化物酶体自噬的报道(Amor et al. 2000;Asakura et al. 2009).细胞自噬在丝状真菌的产孢、程序性细胞死亡、致病及促进细胞生存过程中发挥重要作用.在丝状真菌米曲霉Aspergillus oryzae中,Aoatg8、Aoatg13及Aoatg15的功能缺失将影响其产孢过程,说明自噬在该种真菌的无性生殖中发挥作用(Kikuma & Kitamoto 2011).无独有偶,在其他丝状真菌中,自噬相关基因同样影响着产孢过程,如烟曲霉Aspergillus fumigatus中的Afatg1(Richie et al. 2007),以及稻瘟病菌Magnaporthe oryzae中的Moatg1、Moatg4、Moatg5、Moatg8和Moatg9(Liu et al. 2007)等.稻瘟病的致病过程起始于分生孢子的萌发,随后产生附着胞(appressoria)侵染宿主;附着胞的产生伴随着孢子结构的解体,这一过程需要通过自噬来回收孢子内的物质供给附着胞的生长(Veneault-Fourrey et al. 2006).稻瘟病菌中Moatg1、Moatg2、Moatg3、Moatg4、Moatg5、Moatg6、Moatg7、Moatg8、Moatg9、Moatg10、Moatg12、Moatg13、Moatg15、Moatg16、Moatg17和Moatg18的缺失突变体不能引发稻瘟病,表明这些基因在稻瘟病菌致病过程中发挥作用;这种致病性的缺失是由于附着胞内甘油浓度的下降而导致细胞膨压的降低引起的(Lu et al. 2009).而在大孢粪壳菌Sordaria macrospora中,Smatg8及Smatg4通过选择性及非选择性自噬参与调控营养生长、自噬体的发育及子囊袍子萌发等过程(Oliver & Stefanie 2013). ...
Self-eating and self-killing: crosstalk between autophagy and apoptosis.
1
2007
... 许多丝状真菌对于植物来说是致病菌,农业上,这些真菌引起的真菌病害将导致农作物的减产及质量的下降.传统的防治真菌病害的药物存在着药物残留高,病菌易产生抗药性等问题,而现代农业逐渐强调低毒、低残留、高效及环境相容性.如果可以以真菌自噬为靶点,筛选出特异性抑制真菌自噬的化合物,从而抑制真菌对宿主的侵染,也许可以克服传统农药的弊端.另一方面,许多真菌次级代谢产物具有抗癌抗肿瘤作用.近年来的研究提供了越来越多的证据支持自噬是一种肿瘤抑制途径的观点(Levine 2007).事实上,调节自噬和调节肿瘤发生的信号通路密切重叠,如TOR信号上游抑制子是抑癌基因,包括PTEN、TSC1和TSC2,刺激自噬过程,相反,激活致癌基因TOR的蛋白如Ⅰ类PI3K和Akt等则抑制自噬.尽管目前尚不清楚细胞存活/细胞死亡效应是否与自噬抑制肿瘤的作用有关,但是这些效应可能在癌症治疗过程中十分重要.针对ATG基因用siRNAs特异性的抑制自噬过程往往在这些实验中加速细胞死亡,而不是防止(Maiuri et al. 2007),说明自噬激活代表着一个细胞试图应对细胞毒性药物诱导产生的压力.这表明抑制自噬(而不是激活自噬)可能更有利于癌症治疗.C77,源于Gliocladium sp.的次生代谢产物,属于多硫代二酮哌嗪(epipolythiodioxopiperazines,ETPs)类化合物(Dong et al. 2005;Gardiner et al. 2005),具有广谱的生物活性.本实验室近期的研究结果表明,C77在一定程度上抑制乙肝病毒的复制,而这一过程可能是通过抑制乙肝细胞自噬过程引起的.丝状真菌中的自噬过程影响着次级代谢产物的生成及释放.产黄青霉Penicillium chrysogenum中atg1的缺失突变体在抑制了自噬过程的同时明显提高了青霉素的产量(Bartoszewska et al. 2011b).因而,更好地了解自噬的分子机制将有利于我们以自噬为靶点寻求肿瘤治疗的突破点. ...
Autophagy fights disease through cellular self-digestion.
1
2008
... 根据包裹底物分子及运送至降解细胞器方式的不同,可以将自噬分为巨自噬(macroautophagy),微自噬(microautophagy)和分子伴侣介导的自噬(chaperone-mediated autophagy)等3种(图1),其中巨自噬即通常所指的自噬,在本文此后的部分中,没有特殊标注的“自噬”均代表巨自噬(Mizushima et al. 2008). ...
How to interpret LC3 immunoblotting.
2
2007
... 这一系列的方法具有相对安全、简便,技术及仪器普及性高等优点,一般生化实验室即可达到应用的标准,因而被广泛地用来检测自噬.这一系列的检测方法包括:(1)电子显微镜(Eskelinen et al. 2011),利用电子显微镜分析自噬过程中多种膜结构的连续变化是一种即有效又必要的定性及定量分析方法;(2)Atg8/LC3的Western Blot分析(Mizushima & Yoshimori 2007),Atg8-PE/LC3-II是唯一一种可靠的与自噬体相关联的蛋白标记,通过对其含量的Western Blot分析来间接分析自噬体的含量;(3)荧光显微镜(Kabeya et al. 2000),将绿色荧光蛋白(GFP)或其他荧光蛋白的基因与Atg8/LC3蛋白的基因融合到一起,表达一个具有功能的融合蛋白,可以被应用于间接免疫荧光和荧光显微镜的方法检测自噬. ...
... 自噬过程不仅仅是自噬体的形成和消亡,更重要的是底物分子在整个自噬系统中的传递,即自噬流(autophagic flux).通过检测底物分子在细胞中的传递来分析自噬过程更为恰当.通过自噬流检测自噬的方法包括:(1)自噬性蛋白质的降解,利用放射性氨基酸标记检测细胞内长寿命蛋白的降解来检测自噬过程(Pattingre et al. 2004);(2)LC3-II的周转周期(Tanida et al. 2005),使用蛋白酶抑制剂或改变溶酶体pH的巴弗洛霉素A1阻止溶酶体的降解,或使用阻断溶酶体与自噬体融合的试剂,使得LC3-II的含量不再变化,利用免疫印迹的方法推断LC3-II的周转周期来测量;(3)p62 western印迹(Mizushima & Yoshimori 2007),P62蛋白的作用是联系LC3和泛素化底物,p62被包含到完整的自噬体中,在自噬泡阶段被降解,最近的一项研究表明p62含量的增加与自噬的抑制相关,指出这种蛋白稳定状态下的含量反映了自噬过程的状态;(4)RFP-GFP联合荧光显微镜(Kimura et al. 2007)等. ...
Novel system for monitoring autophagy in the yeast Saccharomyces cerevisiae.
0
1995
Autophagy genes Smatg8 and Smatg4 are required for fruiting-body development, vegetative growth and ascospore germination in the filamentous ascomycete Sordaria macrospora. Autophagy,
1
2013
... 丝状真菌在人类社会中十分常见,在健康、食品、农业等方面都发挥着十分重要的作用,在自然条件及工业发酵条件下,丝状真菌持续经历着营养匮乏,在这种条件下,丝状真菌可能通过自噬来促进细胞存活及分化(Bartoszewska & Kiel 2011a).值得注意的是,到目前为止,对于丝状真菌中选择性自噬途径的研究还不是十分透彻,仅有几篇关于过氧化物酶体自噬的报道(Amor et al. 2000;Asakura et al. 2009).细胞自噬在丝状真菌的产孢、程序性细胞死亡、致病及促进细胞生存过程中发挥重要作用.在丝状真菌米曲霉Aspergillus oryzae中,Aoatg8、Aoatg13及Aoatg15的功能缺失将影响其产孢过程,说明自噬在该种真菌的无性生殖中发挥作用(Kikuma & Kitamoto 2011).无独有偶,在其他丝状真菌中,自噬相关基因同样影响着产孢过程,如烟曲霉Aspergillus fumigatus中的Afatg1(Richie et al. 2007),以及稻瘟病菌Magnaporthe oryzae中的Moatg1、Moatg4、Moatg5、Moatg8和Moatg9(Liu et al. 2007)等.稻瘟病的致病过程起始于分生孢子的萌发,随后产生附着胞(appressoria)侵染宿主;附着胞的产生伴随着孢子结构的解体,这一过程需要通过自噬来回收孢子内的物质供给附着胞的生长(Veneault-Fourrey et al. 2006).稻瘟病菌中Moatg1、Moatg2、Moatg3、Moatg4、Moatg5、Moatg6、Moatg7、Moatg8、Moatg9、Moatg10、Moatg12、Moatg13、Moatg15、Moatg16、Moatg17和Moatg18的缺失突变体不能引发稻瘟病,表明这些基因在稻瘟病菌致病过程中发挥作用;这种致病性的缺失是由于附着胞内甘油浓度的下降而导致细胞膨压的降低引起的(Lu et al. 2009).而在大孢粪壳菌Sordaria macrospora中,Smatg8及Smatg4通过选择性及非选择性自噬参与调控营养生长、自噬体的发育及子囊袍子萌发等过程(Oliver & Stefanie 2013). ...
Analyses of Galpha-interacting protein and activator of G-protein-signaling-3 functions in macroautophagy.
1
2004
... 自噬过程不仅仅是自噬体的形成和消亡,更重要的是底物分子在整个自噬系统中的传递,即自噬流(autophagic flux).通过检测底物分子在细胞中的传递来分析自噬过程更为恰当.通过自噬流检测自噬的方法包括:(1)自噬性蛋白质的降解,利用放射性氨基酸标记检测细胞内长寿命蛋白的降解来检测自噬过程(Pattingre et al. 2004);(2)LC3-II的周转周期(Tanida et al. 2005),使用蛋白酶抑制剂或改变溶酶体pH的巴弗洛霉素A1阻止溶酶体的降解,或使用阻断溶酶体与自噬体融合的试剂,使得LC3-II的含量不再变化,利用免疫印迹的方法推断LC3-II的周转周期来测量;(3)p62 western印迹(Mizushima & Yoshimori 2007),P62蛋白的作用是联系LC3和泛素化底物,p62被包含到完整的自噬体中,在自噬泡阶段被降解,最近的一项研究表明p62含量的增加与自噬的抑制相关,指出这种蛋白稳定状态下的含量反映了自噬过程的状态;(4)RFP-GFP联合荧光显微镜(Kimura et al. 2007)等. ...
Unexpected link between metal ion deficiency and autophagy in Aspergillus fumigatus.
1
2007
... 丝状真菌在人类社会中十分常见,在健康、食品、农业等方面都发挥着十分重要的作用,在自然条件及工业发酵条件下,丝状真菌持续经历着营养匮乏,在这种条件下,丝状真菌可能通过自噬来促进细胞存活及分化(Bartoszewska & Kiel 2011a).值得注意的是,到目前为止,对于丝状真菌中选择性自噬途径的研究还不是十分透彻,仅有几篇关于过氧化物酶体自噬的报道(Amor et al. 2000;Asakura et al. 2009).细胞自噬在丝状真菌的产孢、程序性细胞死亡、致病及促进细胞生存过程中发挥重要作用.在丝状真菌米曲霉Aspergillus oryzae中,Aoatg8、Aoatg13及Aoatg15的功能缺失将影响其产孢过程,说明自噬在该种真菌的无性生殖中发挥作用(Kikuma & Kitamoto 2011).无独有偶,在其他丝状真菌中,自噬相关基因同样影响着产孢过程,如烟曲霉Aspergillus fumigatus中的Afatg1(Richie et al. 2007),以及稻瘟病菌Magnaporthe oryzae中的Moatg1、Moatg4、Moatg5、Moatg8和Moatg9(Liu et al. 2007)等.稻瘟病的致病过程起始于分生孢子的萌发,随后产生附着胞(appressoria)侵染宿主;附着胞的产生伴随着孢子结构的解体,这一过程需要通过自噬来回收孢子内的物质供给附着胞的生长(Veneault-Fourrey et al. 2006).稻瘟病菌中Moatg1、Moatg2、Moatg3、Moatg4、Moatg5、Moatg6、Moatg7、Moatg8、Moatg9、Moatg10、Moatg12、Moatg13、Moatg15、Moatg16、Moatg17和Moatg18的缺失突变体不能引发稻瘟病,表明这些基因在稻瘟病菌致病过程中发挥作用;这种致病性的缺失是由于附着胞内甘油浓度的下降而导致细胞膨压的降低引起的(Lu et al. 2009).而在大孢粪壳菌Sordaria macrospora中,Smatg8及Smatg4通过选择性及非选择性自噬参与调控营养生长、自噬体的发育及子囊袍子萌发等过程(Oliver & Stefanie 2013). ...
Lysosomal turnover, but not a cellular level, of endogenous LC3 is a marker for autophagy.
1
2005
... 自噬过程不仅仅是自噬体的形成和消亡,更重要的是底物分子在整个自噬系统中的传递,即自噬流(autophagic flux).通过检测底物分子在细胞中的传递来分析自噬过程更为恰当.通过自噬流检测自噬的方法包括:(1)自噬性蛋白质的降解,利用放射性氨基酸标记检测细胞内长寿命蛋白的降解来检测自噬过程(Pattingre et al. 2004);(2)LC3-II的周转周期(Tanida et al. 2005),使用蛋白酶抑制剂或改变溶酶体pH的巴弗洛霉素A1阻止溶酶体的降解,或使用阻断溶酶体与自噬体融合的试剂,使得LC3-II的含量不再变化,利用免疫印迹的方法推断LC3-II的周转周期来测量;(3)p62 western印迹(Mizushima & Yoshimori 2007),P62蛋白的作用是联系LC3和泛素化底物,p62被包含到完整的自噬体中,在自噬泡阶段被降解,最近的一项研究表明p62含量的增加与自噬的抑制相关,指出这种蛋白稳定状态下的含量反映了自噬过程的状态;(4)RFP-GFP联合荧光显微镜(Kimura et al. 2007)等. ...
Autophagic fungal cell death is necessary for infection by the rice blast fungus.
1
2006
... 丝状真菌在人类社会中十分常见,在健康、食品、农业等方面都发挥着十分重要的作用,在自然条件及工业发酵条件下,丝状真菌持续经历着营养匮乏,在这种条件下,丝状真菌可能通过自噬来促进细胞存活及分化(Bartoszewska & Kiel 2011a).值得注意的是,到目前为止,对于丝状真菌中选择性自噬途径的研究还不是十分透彻,仅有几篇关于过氧化物酶体自噬的报道(Amor et al. 2000;Asakura et al. 2009).细胞自噬在丝状真菌的产孢、程序性细胞死亡、致病及促进细胞生存过程中发挥重要作用.在丝状真菌米曲霉Aspergillus oryzae中,Aoatg8、Aoatg13及Aoatg15的功能缺失将影响其产孢过程,说明自噬在该种真菌的无性生殖中发挥作用(Kikuma & Kitamoto 2011).无独有偶,在其他丝状真菌中,自噬相关基因同样影响着产孢过程,如烟曲霉Aspergillus fumigatus中的Afatg1(Richie et al. 2007),以及稻瘟病菌Magnaporthe oryzae中的Moatg1、Moatg4、Moatg5、Moatg8和Moatg9(Liu et al. 2007)等.稻瘟病的致病过程起始于分生孢子的萌发,随后产生附着胞(appressoria)侵染宿主;附着胞的产生伴随着孢子结构的解体,这一过程需要通过自噬来回收孢子内的物质供给附着胞的生长(Veneault-Fourrey et al. 2006).稻瘟病菌中Moatg1、Moatg2、Moatg3、Moatg4、Moatg5、Moatg6、Moatg7、Moatg8、Moatg9、Moatg10、Moatg12、Moatg13、Moatg15、Moatg16、Moatg17和Moatg18的缺失突变体不能引发稻瘟病,表明这些基因在稻瘟病菌致病过程中发挥作用;这种致病性的缺失是由于附着胞内甘油浓度的下降而导致细胞膨压的降低引起的(Lu et al. 2009).而在大孢粪壳菌Sordaria macrospora中,Smatg8及Smatg4通过选择性及非选择性自噬参与调控营养生长、自噬体的发育及子囊袍子萌发等过程(Oliver & Stefanie 2013). ...
The molecular mechanism of autophagy.
1
2003
... 自噬是一种广泛存在于真核细胞内的依赖于溶酶体/液泡的降解途径,通过降解细胞内过多或异常的蛋白、细胞器等以维持正常的细胞功能.在自噬过程中,待降解的胞浆大分子物质或细胞器被双层膜泡结构包裹并通过细胞骨架微管网路系统运送至降解细胞器(溶酶体或液泡)中降解(Wang & Klionsky 2003),降解得到的产物分子如氨基酸等被运回细胞质内,重新利用.此外,自噬在某些情况下还能导致细胞死亡,因而被认为是区别于细胞凋亡的另一种细胞程序性死亡形式(Ⅱ型程序性死亡)(Edinger & Thompson 2004),细胞死亡命名委员会(Nomenclature Committee on Cell Death)分别在2009年和2011年讨论并推荐使用“自噬性细胞死亡”(autophagic cell death)这一概念来描述伴随着大量细胞质空泡化,往往(尽管不总是)表示自噬流(autophagic flux)增加的细胞死亡(Kroemer et al. 2009;Galluzzi et al. 2012).“自噬性细胞死亡”这一概念暗示着自噬实际上可以执行细胞消亡,支持这一论证的证据之一是自噬似乎响应于体外选择的化疗药物而介导一些癌细胞的死亡(特别是当缺少重要的细胞凋亡调节因子如BAX和BAK或caspases时)(Grander et al. 2009).同时,自噬作为细胞的管理程序,具有消除有缺陷的蛋白和细胞器,阻止异常蛋白的聚合积累和清除细胞内的病原体的功能,可能是防止衰老、癌症、神经退行性疾病和病原体感染的关键(Levine & Kroemer 2008).已知自噬过程在几乎所有类型细胞的生理学和病理生理学方面都发挥着重要作用,自噬被认为是响应多种刺激的细胞反应(Codogno & Meijer 2005). ...
Autophagosome formation: core machinery and adaptations.
1
2007
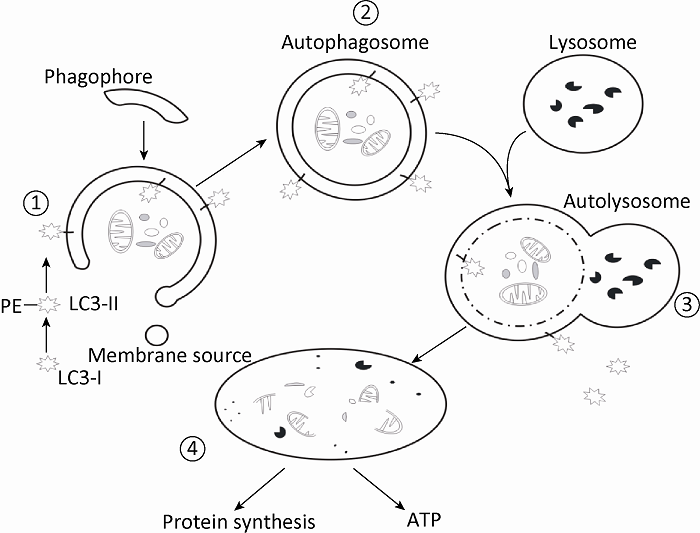
... 以巨自噬为例,自噬的发生过程可人为的划分为4个部分:(1)分隔膜的形成:机体在正常或应激条件下,粗面内质网非核糖体区域、高尔基体等来源的杯状双层膜结构围绕待降解底物;(2)自噬体的形成:随着双层膜结构的延伸和末端融合,待降解底物完全与细胞质隔离,形成自噬体;(3)自噬体的运输和融合:自噬体通过细胞骨架微管网路系统被运输到降解细胞器,紧接着自噬体外层膜与降解细胞器的膜融合,内层膜与底物分子一起进入降解细胞器内部,随后自噬体的内层膜断裂,暴露出底物分子;(4)降解、再循环:底物分子最终被多种水解酶水解成小分子,降解产物被释放回细胞质,循环利用(Xie & Klionsky 2007)(图2).在动物细胞中,与自噬体融合后的溶酶体最后通过再生机制从融合细胞器中再生(Yu et al. 2010). ...
Termination of autophagy and reformation of lysosomes regulated by mTOR.
2
2010
... 以巨自噬为例,自噬的发生过程可人为的划分为4个部分:(1)分隔膜的形成:机体在正常或应激条件下,粗面内质网非核糖体区域、高尔基体等来源的杯状双层膜结构围绕待降解底物;(2)自噬体的形成:随着双层膜结构的延伸和末端融合,待降解底物完全与细胞质隔离,形成自噬体;(3)自噬体的运输和融合:自噬体通过细胞骨架微管网路系统被运输到降解细胞器,紧接着自噬体外层膜与降解细胞器的膜融合,内层膜与底物分子一起进入降解细胞器内部,随后自噬体的内层膜断裂,暴露出底物分子;(4)降解、再循环:底物分子最终被多种水解酶水解成小分子,降解产物被释放回细胞质,循环利用(Xie & Klionsky 2007)(图2).在动物细胞中,与自噬体融合后的溶酶体最后通过再生机制从融合细胞器中再生(Yu et al. 2010). ...
... 丝状真菌在人类社会中十分常见,在健康、食品、农业等方面都发挥着十分重要的作用,在自然条件及工业发酵条件下,丝状真菌持续经历着营养匮乏,在这种条件下,丝状真菌可能通过自噬来促进细胞存活及分化(Bartoszewska & Kiel 2011a).值得注意的是,到目前为止,对于丝状真菌中选择性自噬途径的研究还不是十分透彻,仅有几篇关于过氧化物酶体自噬的报道(Amor et al. 2000;Asakura et al. 2009).细胞自噬在丝状真菌的产孢、程序性细胞死亡、致病及促进细胞生存过程中发挥重要作用.在丝状真菌米曲霉Aspergillus oryzae中,Aoatg8、Aoatg13及Aoatg15的功能缺失将影响其产孢过程,说明自噬在该种真菌的无性生殖中发挥作用(Kikuma & Kitamoto 2011).无独有偶,在其他丝状真菌中,自噬相关基因同样影响着产孢过程,如烟曲霉Aspergillus fumigatus中的Afatg1(Richie et al. 2007),以及稻瘟病菌Magnaporthe oryzae中的Moatg1、Moatg4、Moatg5、Moatg8和Moatg9(Liu et al. 2007)等.稻瘟病的致病过程起始于分生孢子的萌发,随后产生附着胞(appressoria)侵染宿主;附着胞的产生伴随着孢子结构的解体,这一过程需要通过自噬来回收孢子内的物质供给附着胞的生长(Veneault-Fourrey et al. 2006).稻瘟病菌中Moatg1、Moatg2、Moatg3、Moatg4、Moatg5、Moatg6、Moatg7、Moatg8、Moatg9、Moatg10、Moatg12、Moatg13、Moatg15、Moatg16、Moatg17和Moatg18的缺失突变体不能引发稻瘟病,表明这些基因在稻瘟病菌致病过程中发挥作用;这种致病性的缺失是由于附着胞内甘油浓度的下降而导致细胞膨压的降低引起的(Lu et al. 2009).而在大孢粪壳菌Sordaria macrospora中,Smatg8及Smatg4通过选择性及非选择性自噬参与调控营养生长、自噬体的发育及子囊袍子萌发等过程(Oliver & Stefanie 2013). ...





{kind=link}
{kind=link}
{kind=link}
{kind=link}