The cell wall of Aspergillus fumigatus: composition, biosynthesis and function
FANG Wen-Xia1, *, , JIN Cheng1, 2
1 National Engineering Research Centre for Non-Food Biorefinery, Guangxi Academy of Sciences, Nanning, Guangxi 530007, China2 State Key Laboratory of Mycology, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101, China
作者简介:房文霞,房文霞 博士,广西科学院研究员。2009年博士毕业于中国科学院微生物研究所,随后到英国邓迪大学做研究助理,参与了2个英国医学研究理事会的项目。2017年作为高层次人才引进到广西科学院全职工作。多年来利用遗传学、生物化学及结构生物学等手段从事丝状真菌烟曲霉基因的功能研究,细胞壁药物靶标的鉴定及抑制剂的筛选等研究,共计发表SCI论文12篇,包括《Molecular Microbiology》《ACS Chemical Biology》《Microbiology》等第一作者论文6篇。
Aspergillus fumigatus is a ubiquitous saprotrophic filamentous fungus in the environment. It is also the most common and life-threatening serial fungal pathogen, causing a variety of acute or chronic diseases in immunocompromised or immunodeficiency patients, including invasive aspergillosis that with high mortality rate. As a unique extracellular matrix fungal cell wall plays major roles in maintaining cell shape, protecting cells against the external stress, establishing polarized growth, invading new ecological niche and stimulating host immune response. Cell wall components are the biomarkers for the diagnosis of fungal infections, thus cell wall has been considered as the ideal target for the development of antifungal drugs. In recent decades, genetic, biochemical and immunological studies on the cell wall of A. fumigatus have made it an ideal model for studying the fungal cell wall. This review outlines composition, biosynthesis and function of A. fumigatus cell wall, and future research directions are proposed.
传统观念认为细胞壁是一种细胞外惰性骨架结构,然而近年来对机会致病真菌烟曲霉的细胞壁研究表明:细胞壁是个活的动态细胞器。烟曲霉是一种普遍存在于环境中的腐生丝状真菌,同时也是三大机会致病真菌之一,在免疫系统低下或缺陷的人群中可引起多种急慢性疾病。烟曲霉有很强的环境适应性,其细胞壁组分随着细胞周期进程和外部环境的变化而变化,这对理解细胞壁的结构组成和分子合成机制提出了重大挑战。研究表明,烟曲霉细胞壁主要由糖蛋白和多糖组成,其中多糖含量高达90%(Fontaine et al. 2000)。鉴定细胞壁各组分使用方法是碱处理细胞壁后,将碱可溶与碱不溶组分分离,再用酶解或酸水解的方法处理两种组分,最后用经典的糖分析鉴定方法测定单糖或多糖的含量,最终确定细胞壁的组分及组成(Fontaine et al. 2000)。
1 烟曲霉细胞壁的组分和合成
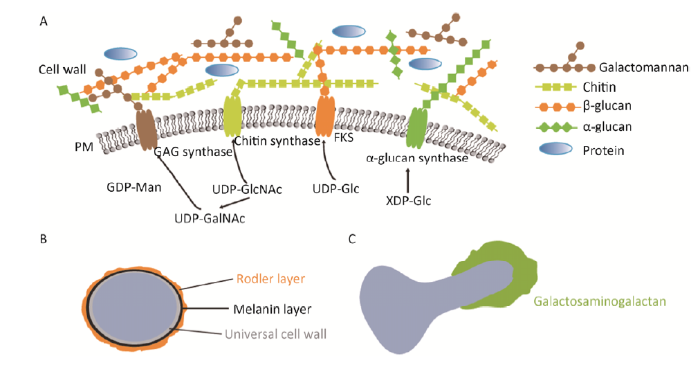
法国巴斯德研究所曲霉中心Jean-Paul Latgé实验室30多年来一直致力于烟曲霉细胞壁的研究。经过多年来的努力目前确定烟曲霉细胞壁主要组分为碱不溶性的纤维状骨架,包括几丁质、甘露聚糖、β-1,3-葡聚糖以及β-1,6-分支的β-1,3-葡聚糖,包埋于无定形的碱可溶组分中包括α-1,3-葡聚糖和半乳甘露聚糖(Fontaine et al. 2000)。烟曲霉细胞壁约占细胞干重的30%,各多糖组分含量大约为7%-15%的几丁质、20%-35%的β-1,3-葡聚糖、35%-46%的α-1,3-葡聚糖和20%-25%的半乳甘露聚糖(Gastebois et al. 2009)。多糖的生物合成起始于胞内活化的核苷酸糖,参与核苷酸糖合成的酶为潜在的药物靶标(Fang et al. 2013a,2013b;Fang 2017)。这些核苷酸糖在一系列跨膜合成酶、转糖苷酶和糖基水解酶的作用下构建成兼具坚固性和柔韧性的细胞壁。三维网络状结构的细胞壁一直处于动态变化中,其组分随着形态发生而发生改变,如休眠期的孢子具有黑色素和rodlet输水蛋白层,在营养菌丝生长过程中这些被逐渐降解,同时出现菌丝特有组分半乳糖氨基半乳聚糖(GAG)(Fontaine et al. 2011;Briard et al. 2016)(图1)。
β-1,3-葡聚糖的合成以细胞内葡萄糖的活化形式UDP-Glc为合成前体,由跨膜葡聚糖合成酶复合体催化合成。早在1993年Latgé实验室就利用放射性标记的UDP(14C)-glucose从烟曲霉菌丝体膜组分中提取到了葡聚糖合成酶复合体,发现其由催化亚基和调控亚基组成(Beauvais et al. 2001)。催化亚基为具有16个跨膜结构域的膜蛋白,是棘白菌素类药物的靶标(Douglas et al. 1997)。催化亚基由基因fks1编码,在烟曲霉基因组中仅有一个拷贝。最早的研究表明 fks1是重要基因(Firon et al. 2002;Mouyna et al. 2004),然而Dichtl et al.(2015)最近发现fks1不是必需的,但其缺失突变株的菌落生长非常缓慢,表现为细胞壁β-1,3-葡聚糖缺失伴随着几丁质和半乳糖氨基半乳聚糖的补偿性增加以及甘露聚糖含量的显著下降。这可能是棘白菌素类药物只能抑制但不能杀死烟曲霉的原因。烟曲霉Rho-GTPase Rho1被鉴定为FKS1的调控亚基(Beauvais et al. 2001),遗传学实验表明它也是必需基因,缺失后导致细胞壁完整性途径(CWI)受损,细胞质渗漏裂解及最终不能存活(Dichtl et al. 2012)。
1.2 几丁质的生物合成
几丁质是β-1,4-连接的N-乙酰葡萄糖胺线性聚合而成。已知几丁质由跨膜几丁质合成酶(Chitin synthase,CHS)以UDP- N-乙酰葡萄糖胺为前体合成。但几丁质合成机理尚不清楚,原因之一是丝状真菌中 CHS基因数量较多(Muszkieta et al. 2014)。基于生物信息学分析烟曲霉有8个chs基因分属于两个家族。家族一的chsA、chsB和chsC对几丁质的合成贡献不大,但chsG单突变株及与chsA/B/C的多重突变株有分生孢子减少,菌丝形态改变、生长下降和几丁质合成酶活性降低等表型(Mellado et al. 1996;Mellado et al. 2003)。家族二包括csmA、csmB、chsF和chsD 4个基因,4重敲除株表现出菌丝形态和孢子的变化,但几丁质含量几乎不受影响。敲除两个家族的4个基因(ΔcsmA/ΔcsmB/ ΔchsF/ΔchsG)后菌丝生长非常缓慢(Muszkieta et al. 2014),类似于fks1突变株,表明两个家族的几丁质合成酶共同协作参与烟曲霉几丁质的合成,也表明几丁质的合成对菌丝生长的重要性。构巢曲霉的细胞壁结构和烟曲霉的非常相似,但其几丁质合成酶的分类和功能与烟曲霉不同(Fujiwara et al. 2000;Tsuizaki et al. 2009),说明不同真菌中几丁质合成酶的功能根据细胞壁整体合成调控的需求有所不同。
1.3 α-1,3-葡聚糖的合成
α-1,3-葡聚糖是细胞壁的无定形组分,约占细胞壁含量的40%。烟曲霉有3种α-1,3-葡聚糖合成酶(AGS1、AGS2和AGS3),都是高分子量的跨膜酶 (>200kDa),包括一个淀粉酶样的结构域和一个似糖原合成酶样的结构域。3个基因在菌丝生长期均为高表达但都不是必需基因(Beauvais et al. 2005;Maubon et al. 2006;Henry et al. 2012)。三重缺失株不含α-1,3-葡聚糖但菌丝营养生长不受影响,菌丝的形态通过β-1,3-葡聚糖和几丁质含量的增加来维持(Henry et al. 2012)。目前对α-1,3-葡聚糖合成酶的底物和体外合成活性尚不清楚。
1.4 半乳甘露聚糖的合成
半乳甘露聚糖的合成需要甘露糖基转移酶和半乳糖基转移酶的协同作用,组装线性的α-甘露聚糖骨架和β-1,5-连接的半乳呋喃糖残基短链。烟曲霉的甘露聚糖合成以GDP-甘露糖为底物,由甘露糖基转移酶催化合成。每个甘露聚糖骨架平均有50个甘露糖残基,由多个α-连接的-M2-M2-M6- M2-重复单元组成(Latge et al. 1994),与葡聚糖-几丁质核心骨架共价连接。目前发现烟曲霉有11个甘露糖基转移酶负责α-1,6-和α-1,2-甘露糖苷键连接,但完全敲除这11个酶并没有导致菌丝体细胞壁中甘露聚糖含量的降低,仅分生孢子细胞壁的甘露聚糖含量减少(Henry et al. 2016)。烟曲霉半乳甘露聚糖的半乳糖以半乳呋喃糖(Glaf)的形式存在,这种糖在许多致病菌和寄生虫中是很重要的己糖构型方式(Latge et al. 1994)。UDP-Galf是半乳甘露聚糖的合成前体(Lamarre et al. 2009),由UDP-半乳吡喃糖在UDP-半乳糖变位酶的催化下转化而成。半乳甘露聚糖的合成最初是在内质网中进行,随后在高尔基复合体中进一步延伸并共价结合β-1,3-葡聚糖(Engel et al. 2012)。
1.5 半乳糖氨基半乳聚糖的合成
另一种含半乳糖的聚合物是半乳糖氨基半乳聚糖(GAG),是只在营养菌丝体中发现的表面多糖。这种糖聚合物呈线性不溶于水,平均大小为100kDa,由半乳吡喃糖,半乳糖胺和N-乙酰半乳糖胺(GalNAc)以α-1,4 -糖苷键连接而成,并沿着多糖链随机分布(Fontaine et al. 2011)。与其他糖聚合物一样,GAG的合成使用UDP-半乳糖和UDP-GalNAc为底物(Briard et al. 2016),并推测出几个与GAG合成相关的酶(Bamford et al. 2015;Gravelat et al. 2013)。最近蛋白质拓扑结构预测显示CAZy家族 (http://www.cazy.org)GT4 的一种α-糖基转移酶有14个跨质膜结构域,可能是GAG合成酶(Briard et al. 2016)。然而目前尚没有GAG合成酶的体外活性证明,这种酶是如何识别并连接GAG中不同的糖仍然是个谜。
2 细胞壁多糖合成后的修饰与组装
新合成的多糖链在细胞壁空间需要特定糖基转移酶和水解酶的共同作用进行重构、交联以产生细胞壁的三维网络结构,其中以β-1,3-葡聚糖的修饰最为多样。由葡聚糖合成酶复合体合成的线性β-1,3-葡聚糖需要经过延伸、分支和水解等过程才构成细胞骨架。葡聚糖的延伸是由糖基转移酶GEL家族负责,将水解后糖链的还原端转运到另一个葡聚糖分子的非还原端,从而延伸葡聚糖链。GEL属于CAZy数据库中的GH72家族,在烟曲霉中有7个成员,其中GEL4的缺失是致死的(Mouyna et al. 2005;Gastebois et al. 2010a),其他成员对葡聚糖的合成、形态发生、细胞壁完整性途径和致病力有不同程度的影响(Mouyna et al. 2005;Zhao et al. 2014)。最近的研究表明含有碳水化合物结合域(carbohydrate binding module,CBM)的GEL4除了延伸β-1,3-葡聚糖外还可以建立分支,具有双重功能(Aimanianda et al. 2017)。BGT1和BGT2基因也具有β-1,3-葡聚糖的分支功能,可以产生β-1,6-分支的β-1,3-葡聚糖,但它们的缺失没有引起细胞壁表型(Gastebois et al. 2010b),说明在细胞壁组装中它们并非重要基因。孢子萌发或菌丝分支生长中需要葡萄糖苷酶水解β-1,3-葡聚糖以增加细胞壁的可塑性。烟曲霉基因组含有多个β-1,3-葡聚糖酶,目前对其中eng1和eng2基因的功能分析发现它们的缺失未导致任何细胞壁缺陷(Mouyna et al. 2002;Hartl et al. 2011)。
几丁质作为提供坚固性的细胞壁组分在形态发生中一直处于动态变化。几丁质酶水解β-1,4-糖苷键从而分解几丁质。烟曲霉有5个植物型、12个细菌型几丁质酶(Taib et al. 2005)。植物型几丁质酶的五重敲除株与细菌型中AfChiB1的缺失都没有明显的表型(Jaques et al. 2003;Alcazar-Fuoli et al. 2011),说明不同类型的几丁质酶功能可以互补。另外几丁质脱乙酰酶和α-1,3-葡聚糖水解酶等在烟曲霉中也有多个冗余基因,利用遗传学技术分析这些酶的功能尚在进行中。
细胞壁多糖到达细胞壁空间后的最后一步是各个组分之间的交联。行驶交联功能的酶通常是通过GPI锚定在细胞壁上的蛋白。酵母中发现CRH家族负责几丁质与葡聚糖之间的交联,CRH缺失后的突变株表现出子细胞与母细胞连接处形态的异常(Cabib et al. 2007;Blanco et al. 2012)。烟曲霉CRH家族有5个成员,我们最近的实验结果表明:整个家族的五重突变株未见任何菌丝体或分生孢子表型(未发表文章),让人开始质疑 β-葡聚糖-几丁质交联在烟曲霉细胞壁结构化中的作用。
3 细胞壁的动态变化
烟曲霉有3种细胞形态:营养菌丝体形态,主要分解土壤中的有机物质维持碳氮循环;无性分生孢子形态,主要介导在空气中的传播;休眠的囊孢子状态,确保自身的长期生存。分生孢子和菌丝体的细胞壁结构组分非常相似,但两种形态又有各自特有的细胞壁外层组分(图1B、图1C)(Latge & Beauvais 2014)。分生孢子的外层有黑色素层,且被rodlet层覆盖。这个rodlet层完全是由 roda基因编码的疏水蛋白构成,具有8个半胱氨酸残基形成的4个二硫键排列成淀粉样从而使分生孢子细胞壁疏水(Aimanianda et al. 2009)。位于rodlet层下面的是二羟基萘(DHN)黑色素层。基因敲除实验表明黑色素决定了孢子细胞壁的结构和硬度(Heinekamp et al. 2012;Bayry et al. 2014)。最近Latgé实验室构建的ags1-3、roda和pks五重缺失株产生白色亲水性分生孢子且数量急剧减少,说明孢子外层的这些组分与产孢能力密切相关。五重缺失株的菌丝体生长不受影响,表明碱不溶性多糖组分是菌丝体细胞壁必需的(Latge et al. 2017)。
分生孢子在萌发时首先是经过膨胀阶段,此时rodlet层被天冬氨酸蛋白酶降解,疏水性孢子开始变成亲水性的。随着细胞内渗透压的增加孢子开始膨大,细胞壁多糖被糖基水解酶降解并伴有新的多糖层开始合成。膨胀的孢子转向极性生长时芽管开始出现,此时黑色素层被瓦解,膨胀期合成的多糖层构成了菌丝的细胞壁,随后有细胞外基质成分GAG覆盖于新的细胞壁表面(Gravelat et al. 2013)。
4 细胞壁组分的功能
细胞壁对真菌的生长和抵抗环境压力至关重要,同时它也是病原真菌与宿主免疫细胞最先接触的细胞器。先天性免疫反应对于有效及时地识别烟曲霉并诱导炎症网络清除非常重要(Becker et al. 2015)。在免疫活性宿主中,肺泡巨噬细胞(AM)和嗜中性粒细胞每天需要清除数百至上千的烟曲霉分生孢子以防止肺部的炎性损伤(Underhill et al. 2015)。肺泡巨噬细胞(alveolar macrophages,AM)是唯一可以进入肺泡腔的吞噬细胞而且是第一个接触吸入孢子的细胞。嗜中性粒细胞是烟曲霉的必需免疫效应细胞,因为严重嗜中性白血球减少症是侵袭性曲霉病发展的主要危险因素(Mircescu et al. 2009)。微小的烟曲霉孢子被吸入宿主肺部时,细胞壁组分有的发挥激活宿主免疫反应的作用,有的则保护孢子并促进其在宿主中的定殖和生存(表1)。
Table 1
表1
表1 烟曲霉细胞壁主要组分及其功能
Table 1 Aspergillus fumigatus cell wall components and functions
细胞壁组分 Cell wall components
功能 Function
几丁质 Chitin
细胞壁主要骨架组分,提供硬度和坚固性 Cell wall skeletal structure for hardness and rigidity 可能激活宿主免疫反应 May activate host immune response
β-1,3-葡聚糖 β-1,3-glucan
细胞壁主要骨架组分,提供硬度和坚固性 Cell wall skeletal structure for hardness and rigidity 细胞壁PAMPs之一,能被宿主Dectin-1识别,激活宿主免疫反应 Cell wall PAMP recognized by dectin-1, activating host immune response
α-1,3-葡聚糖 α-1,3-glucan
细胞壁无定形结构,缺失后不影响细胞生存但致病力丧失 Cell wall amorphous structure, viable but avirulent after abolishment 与吞噬体发生相关,遮蔽PAMPs,减少细胞壁通透性,激活宿主免疫反应,也可能为PAMPs Associated with phagosome biogenesis, masking cell wall PAMPs, reducing cell wall permeability, activating host immune response, one of cell wall PAMPs
疏水棒状小体 Rodlets
孢子特有组分,使孢子成疏水性,利于孢子扩散 Unique conidia component, protecting conidia against dehydration, favoring conidial dispersion 屏蔽PAMPs,阻止先天性免疫反应的启动,促进真菌在宿主中的定殖和生存 Masking PAMPs, preventing the activation of innate immunity response, favoring growth in host
黑色素 Melanin
孢子特有组分,保护孢子不被UV辐射 Unique conidia component, protecting conidia against UV radiation 屏蔽PAMPs,阻止吞噬体发生和酸化,抑制抗真菌ROS的产生,抑制LC3相关吞噬作用,促进真菌在宿主中的生长 Masking PAMPs, blocking phagosome biogenesis and acidification, sequestration of ROS, inhibition of LC3-associated phagocytosis, favoring growth in host
半乳甘露聚糖 Galactomannan
细胞壁主要骨架组分 Cell wall skeletal structure 细胞壁PAMPs之一,被宿主Dectin-2识别,激活宿主免疫反应 Cell wall PAMP recognized by dectin-2, activating the immune response of the host
半乳糖氨基半乳聚糖 Galactosaminogalactan
菌丝特有组分,参与生物膜的形成 Unique component in hyphae, involved in biofilm formation 粘附宿主细胞,诱导中性粒细胞凋亡,屏蔽β-glucan组分,激活血小板,促进真菌在宿主中的生长 Adhesion to host cells, induction of neutrophil apoptosis, masking β-glucan, activation of platelet, favoring growth in host
细胞壁含有病原体相关分子模式(pathogen- associated molecular patterns,PAMPs),可被宿主先天免疫细胞的模式识别受体(pattern recognition receptors,PRR)识别,这对免疫反应的启动是必不可少的。目前发现的PRR包括Toll样受体(TLRs),C-型凝集素受体(CLRs),NOD样受体(NLRs)和可溶性PRRs。研究结果表明:细胞壁的β-1,3-葡聚糖可以被CLRs中的Dectin-1特异性识别(Herre et al. 2004),从而激活了Dectin-1信号通路(Gersuk et al. 2006)和PRRs调节吞噬体成熟的能力,初步揭示了真菌利用PAMPs逃离巨噬细胞杀伤的机制(Mansour et al. 2013)。
几丁质长期以来在过敏和直接抗真菌响应方面发挥着重要作用。对几丁质的免疫应答是细胞类型特异性的,并随着几丁质颗粒的浓度和大小的不同而不同,在人和小鼠中有不一致的报道(Wagener et al. 2014)。几丁质能诱导人几丁三糖糖苷酶产生,降解不溶性几丁质为可溶性壳寡糖,从而作为更强的免疫刺激剂(Wiesner et al. 2015)激活宿主免疫反应。在小鼠中,几丁质选择性诱导IL-10产生但不刺激促炎细胞因子(Wagener et al. 2014)的产生。不同宿主中免疫应答结果不一致的一个原因是目前没有几丁质与受体直接结合的证据。过去发现的几种几丁质受体(Levitz et al. 2015)诱导产生细胞因子的作用仍然存在争议。
α-1,3-葡聚糖可能与吞噬体生物发生相关,因为AGS三重缺失株孢子细胞壁表面的PAMPs容易暴露出来,被吞噬体识别并清除,同时细胞壁对宿主效应分子的通透性增加,最终使三重突变株几乎无感染毒力(Beauvais et al. 2013)。
GAG是菌丝特有的组分,参与生物膜的形成(Fontaine et al. 2011),能促进菌丝与巨噬细胞、嗜中性粒细胞和血小板的结合(Rambach et al. 2015)。另外还具有以下的直接免疫抑制作用:(1)通过NK细胞依赖性机制引发多形核中性粒细胞(polymorphonuclear neutrophil,PMN)凋亡并抑制PMN趋化性(Robinet et al. 2014);(2)介导对依赖于NADPH氧化酶的嗜中性粒细胞细胞外捕获的抗性(Lee et al. 2015)。GM在菌丝生长过程中被分泌出来,是用于IA早期诊断的有效抗原之一(Foy et al. 2007)。Bio-Rad开发的基于GM单克隆抗体的PlateliaTM检测试剂盒因其较好的特异性和灵敏性被广泛用于临床检测中。最近有报道称,Dectin-2与GM结合(Reedy et al. 2016),但这种C型凝集素的免疫功能尚不完全清楚。
Deletion of the alpha-(1,3)- glucan synthase genes induces a restructuring of the conidial cell wall responsible for the avirulence of Aspergillus fumigatus.
Characterization of essential genes by parasexual genetics in the human fungal pathogen Aspergillus fumigatus: impact of genomic rearrangements associated with electroporation of DNA.
Evidence that the Aspergillus nidulans class I and class II chitin synthase genes, chsC and chsA, share critical roles in hyphal wall integrity and conidiophore development.
Deciphering the role of the chitin synthase families 1 and 2 in the in vivo and in vitro growth of Aspergillus fumigatus by multiple gene targeting deletion.
Surface hydrophobin prevents immune recognition of airborne fungal spores
2
2009
... 烟曲霉有3种细胞形态:营养菌丝体形态,主要分解土壤中的有机物质维持碳氮循环;无性分生孢子形态,主要介导在空气中的传播;休眠的囊孢子状态,确保自身的长期生存.分生孢子和菌丝体的细胞壁结构组分非常相似,但两种形态又有各自特有的细胞壁外层组分(图1B、图1C)(Latge & Beauvais 2014).分生孢子的外层有黑色素层,且被rodlet层覆盖.这个rodlet层完全是由 roda基因编码的疏水蛋白构成,具有8个半胱氨酸残基形成的4个二硫键排列成淀粉样从而使分生孢子细胞壁疏水(Aimanianda et al. 2009).位于rodlet层下面的是二羟基萘(DHN)黑色素层.基因敲除实验表明黑色素决定了孢子细胞壁的结构和硬度(Heinekamp et al. 2012;Bayry et al. 2014).最近Latgé实验室构建的ags1-3、roda和pks五重缺失株产生白色亲水性分生孢子且数量急剧减少,说明孢子外层的这些组分与产孢能力密切相关.五重缺失株的菌丝体生长不受影响,表明碱不溶性多糖组分是菌丝体细胞壁必需的(Latge et al. 2017). ...
... 作为孢子特有组分的rodlet层不仅有利于孢子的扩散,同时阻止细胞壁暴露PAMPs从而不能快速诱导先天性免疫反应(Aimanianda et al. 2009).孢子表面的黑色素具有多重功能,不仅保护孢子不被UV辐射,在病理发生时屏蔽细胞壁PAMPs,而且有很强的抗AM吞噬功能.目前发现的黑色素抗吞噬的主要机制为:(1)阻止吞噬体的酸化(Amin et al. 2014);(2)抑制负责产生抗真菌ROS的NADPH氧化酶(Akoumianaki et al. 2016);(3)阻断LC3相关吞噬作用的特殊自噬途径,避免其被吞噬(Akoumianaki et al. 2016). ...
The dual activity responsible for the elongation and branching of beta-(1,3)-glucan in the fungal cell wall
1
2017
... 新合成的多糖链在细胞壁空间需要特定糖基转移酶和水解酶的共同作用进行重构、交联以产生细胞壁的三维网络结构,其中以β-1,3-葡聚糖的修饰最为多样.由葡聚糖合成酶复合体合成的线性β-1,3-葡聚糖需要经过延伸、分支和水解等过程才构成细胞骨架.葡聚糖的延伸是由糖基转移酶GEL家族负责,将水解后糖链的还原端转运到另一个葡聚糖分子的非还原端,从而延伸葡聚糖链.GEL属于CAZy数据库中的GH72家族,在烟曲霉中有7个成员,其中GEL4的缺失是致死的(Mouyna et al. 2005;Gastebois et al. 2010a),其他成员对葡聚糖的合成、形态发生、细胞壁完整性途径和致病力有不同程度的影响(Mouyna et al. 2005;Zhao et al. 2014).最近的研究表明含有碳水化合物结合域(carbohydrate binding module,CBM)的GEL4除了延伸β-1,3-葡聚糖外还可以建立分支,具有双重功能(Aimanianda et al. 2017).BGT1和BGT2基因也具有β-1,3-葡聚糖的分支功能,可以产生β-1,6-分支的β-1,3-葡聚糖,但它们的缺失没有引起细胞壁表型(Gastebois et al. 2010b),说明在细胞壁组装中它们并非重要基因.孢子萌发或菌丝分支生长中需要葡萄糖苷酶水解β-1,3-葡聚糖以增加细胞壁的可塑性.烟曲霉基因组含有多个β-1,3-葡聚糖酶,目前对其中eng1和eng2基因的功能分析发现它们的缺失未导致任何细胞壁缺陷(Mouyna et al. 2002;Hartl et al. 2011). ...
Aspergillus cell wall melanin blocks LC3-associated phagocytosis to promote pathogenicity
2
2016
... 作为孢子特有组分的rodlet层不仅有利于孢子的扩散,同时阻止细胞壁暴露PAMPs从而不能快速诱导先天性免疫反应(Aimanianda et al. 2009).孢子表面的黑色素具有多重功能,不仅保护孢子不被UV辐射,在病理发生时屏蔽细胞壁PAMPs,而且有很强的抗AM吞噬功能.目前发现的黑色素抗吞噬的主要机制为:(1)阻止吞噬体的酸化(Amin et al. 2014);(2)抑制负责产生抗真菌ROS的NADPH氧化酶(Akoumianaki et al. 2016);(3)阻断LC3相关吞噬作用的特殊自噬途径,避免其被吞噬(Akoumianaki et al. 2016). ...
... );(3)阻断LC3相关吞噬作用的特殊自噬途径,避免其被吞噬(Akoumianaki et al. 2016). ...
Functional analysis of the fungal/plant class chitinase family in Aspergillus fumigatus.
1
2011
... 几丁质作为提供坚固性的细胞壁组分在形态发生中一直处于动态变化.几丁质酶水解β-1,4-糖苷键从而分解几丁质.烟曲霉有5个植物型、12个细菌型几丁质酶(Taib et al. 2005).植物型几丁质酶的五重敲除株与细菌型中AfChiB1的缺失都没有明显的表型(Jaques et al. 2003;Alcazar-Fuoli et al. 2011),说明不同类型的几丁质酶功能可以互补.另外几丁质脱乙酰酶和α-1,3-葡聚糖水解酶等在烟曲霉中也有多个冗余基因,利用遗传学技术分析这些酶的功能尚在进行中. ...
Melanin dependent survival of Apergillus fumigatus conidia in lung epithelial cells.
1
2014
... 作为孢子特有组分的rodlet层不仅有利于孢子的扩散,同时阻止细胞壁暴露PAMPs从而不能快速诱导先天性免疫反应(Aimanianda et al. 2009).孢子表面的黑色素具有多重功能,不仅保护孢子不被UV辐射,在病理发生时屏蔽细胞壁PAMPs,而且有很强的抗AM吞噬功能.目前发现的黑色素抗吞噬的主要机制为:(1)阻止吞噬体的酸化(Amin et al. 2014);(2)抑制负责产生抗真菌ROS的NADPH氧化酶(Akoumianaki et al. 2016);(3)阻断LC3相关吞噬作用的特殊自噬途径,避免其被吞噬(Akoumianaki et al. 2016). ...
Sph3 is a glycoside hydrolase required for the biosynthesis of galactosaminogalactan in Aspergillus fumigatus.
1
2015
... 另一种含半乳糖的聚合物是半乳糖氨基半乳聚糖(GAG),是只在营养菌丝体中发现的表面多糖.这种糖聚合物呈线性不溶于水,平均大小为100kDa,由半乳吡喃糖,半乳糖胺和N-乙酰半乳糖胺(GalNAc)以α-1,4 -糖苷键连接而成,并沿着多糖链随机分布(Fontaine et al. 2011).与其他糖聚合物一样,GAG的合成使用UDP-半乳糖和UDP-GalNAc为底物(Briard et al. 2016),并推测出几个与GAG合成相关的酶(Bamford et al. 2015;Gravelat et al. 2013).最近蛋白质拓扑结构预测显示CAZy家族 (http://www.cazy.org)GT4 的一种α-糖基转移酶有14个跨质膜结构域,可能是GAG合成酶(Briard et al. 2016).然而目前尚没有GAG合成酶的体外活性证明,这种酶是如何识别并连接GAG中不同的糖仍然是个谜. ...
Surface structure characterization of Aspergillus fumigatus conidia mutated in the melanin synthesis pathway and their human cellular immune response.
1
2014
... 烟曲霉有3种细胞形态:营养菌丝体形态,主要分解土壤中的有机物质维持碳氮循环;无性分生孢子形态,主要介导在空气中的传播;休眠的囊孢子状态,确保自身的长期生存.分生孢子和菌丝体的细胞壁结构组分非常相似,但两种形态又有各自特有的细胞壁外层组分(图1B、图1C)(Latge & Beauvais 2014).分生孢子的外层有黑色素层,且被rodlet层覆盖.这个rodlet层完全是由 roda基因编码的疏水蛋白构成,具有8个半胱氨酸残基形成的4个二硫键排列成淀粉样从而使分生孢子细胞壁疏水(Aimanianda et al. 2009).位于rodlet层下面的是二羟基萘(DHN)黑色素层.基因敲除实验表明黑色素决定了孢子细胞壁的结构和硬度(Heinekamp et al. 2012;Bayry et al. 2014).最近Latgé实验室构建的ags1-3、roda和pks五重缺失株产生白色亲水性分生孢子且数量急剧减少,说明孢子外层的这些组分与产孢能力密切相关.五重缺失株的菌丝体生长不受影响,表明碱不溶性多糖组分是菌丝体细胞壁必需的(Latge et al. 2017). ...
Deletion of the alpha-(1,3)- glucan synthase genes induces a restructuring of the conidial cell wall responsible for the avirulence of Aspergillus fumigatus.
1
2013
... α-1,3-葡聚糖可能与吞噬体生物发生相关,因为AGS三重缺失株孢子细胞壁表面的PAMPs容易暴露出来,被吞噬体识别并清除,同时细胞壁对宿主效应分子的通透性增加,最终使三重突变株几乎无感染毒力(Beauvais et al. 2013). ...
Glucan synthase complex of Aspergillus fumigatus.
2
2001
... β-1,3-葡聚糖的合成以细胞内葡萄糖的活化形式UDP-Glc为合成前体,由跨膜葡聚糖合成酶复合体催化合成.早在1993年Latgé实验室就利用放射性标记的UDP(14C)-glucose从烟曲霉菌丝体膜组分中提取到了葡聚糖合成酶复合体,发现其由催化亚基和调控亚基组成(Beauvais et al. 2001).催化亚基为具有16个跨膜结构域的膜蛋白,是棘白菌素类药物的靶标(Douglas et al. 1997).催化亚基由基因fks1编码,在烟曲霉基因组中仅有一个拷贝.最早的研究表明 fks1是重要基因(Firon et al. 2002;Mouyna et al. 2004),然而Dichtl et al.(2015)最近发现fks1不是必需的,但其缺失突变株的菌落生长非常缓慢,表现为细胞壁β-1,3-葡聚糖缺失伴随着几丁质和半乳糖氨基半乳聚糖的补偿性增加以及甘露聚糖含量的显著下降.这可能是棘白菌素类药物只能抑制但不能杀死烟曲霉的原因.烟曲霉Rho-GTPase Rho1被鉴定为FKS1的调控亚基(Beauvais et al. 2001),遗传学实验表明它也是必需基因,缺失后导致细胞壁完整性途径(CWI)受损,细胞质渗漏裂解及最终不能存活(Dichtl et al. 2012). ...
... 不是必需的,但其缺失突变株的菌落生长非常缓慢,表现为细胞壁β-1,3-葡聚糖缺失伴随着几丁质和半乳糖氨基半乳聚糖的补偿性增加以及甘露聚糖含量的显著下降.这可能是棘白菌素类药物只能抑制但不能杀死烟曲霉的原因.烟曲霉Rho-GTPase Rho1被鉴定为FKS1的调控亚基(Beauvais et al. 2001),遗传学实验表明它也是必需基因,缺失后导致细胞壁完整性途径(CWI)受损,细胞质渗漏裂解及最终不能存活(Dichtl et al. 2012). ...
Two alpha(1-3) glucan synthases with different functions in Aspergillus fumigatus.
1
2005
... α-1,3-葡聚糖是细胞壁的无定形组分,约占细胞壁含量的40%.烟曲霉有3种α-1,3-葡聚糖合成酶(AGS1、AGS2和AGS3),都是高分子量的跨膜酶 (>200kDa),包括一个淀粉酶样的结构域和一个似糖原合成酶样的结构域.3个基因在菌丝生长期均为高表达但都不是必需基因(Beauvais et al. 2005;Maubon et al. 2006;Henry et al. 2012).三重缺失株不含α-1,3-葡聚糖但菌丝营养生长不受影响,菌丝的形态通过β-1,3-葡聚糖和几丁质含量的增加来维持(Henry et al. 2012).目前对α-1,3-葡聚糖合成酶的底物和体外合成活性尚不清楚. ...
Antifungal innate immunity: recognition and inflammatory networks
1
2015
... 细胞壁对真菌的生长和抵抗环境压力至关重要,同时它也是病原真菌与宿主免疫细胞最先接触的细胞器.先天性免疫反应对于有效及时地识别烟曲霉并诱导炎症网络清除非常重要(Becker et al. 2015).在免疫活性宿主中,肺泡巨噬细胞(AM)和嗜中性粒细胞每天需要清除数百至上千的烟曲霉分生孢子以防止肺部的炎性损伤(Underhill et al. 2015).肺泡巨噬细胞(alveolar macrophages,AM)是唯一可以进入肺泡腔的吞噬细胞而且是第一个接触吸入孢子的细胞.嗜中性粒细胞是烟曲霉的必需免疫效应细胞,因为严重嗜中性白血球减少症是侵袭性曲霉病发展的主要危险因素(Mircescu et al. 2009).微小的烟曲霉孢子被吸入宿主肺部时,细胞壁组分有的发挥激活宿主免疫反应的作用,有的则保护孢子并促进其在宿主中的定殖和生存(表1). ...
Crosslinks in the cell wall of budding yeast control morphogenesis at the mother-bud neck
1
2012
... 细胞壁多糖到达细胞壁空间后的最后一步是各个组分之间的交联.行驶交联功能的酶通常是通过GPI锚定在细胞壁上的蛋白.酵母中发现CRH家族负责几丁质与葡聚糖之间的交联,CRH缺失后的突变株表现出子细胞与母细胞连接处形态的异常(Cabib et al. 2007;Blanco et al. 2012).烟曲霉CRH家族有5个成员,我们最近的实验结果表明:整个家族的五重突变株未见任何菌丝体或分生孢子表型(未发表文章),让人开始质疑 β-葡聚糖-几丁质交联在烟曲霉细胞壁结构化中的作用. ...
Galactosaminogalactan of Aspergillus fumigatus, a bioactive fungal polymer.
3
2016
... 法国巴斯德研究所曲霉中心Jean-Paul Latgé实验室30多年来一直致力于烟曲霉细胞壁的研究.经过多年来的努力目前确定烟曲霉细胞壁主要组分为碱不溶性的纤维状骨架,包括几丁质、甘露聚糖、β-1,3-葡聚糖以及β-1,6-分支的β-1,3-葡聚糖,包埋于无定形的碱可溶组分中包括α-1,3-葡聚糖和半乳甘露聚糖(Fontaine et al. 2000).烟曲霉细胞壁约占细胞干重的30%,各多糖组分含量大约为7%-15%的几丁质、20%-35%的β-1,3-葡聚糖、35%-46%的α-1,3-葡聚糖和20%-25%的半乳甘露聚糖(Gastebois et al. 2009).多糖的生物合成起始于胞内活化的核苷酸糖,参与核苷酸糖合成的酶为潜在的药物靶标(Fang et al. 2013a,2013b;Fang 2017).这些核苷酸糖在一系列跨膜合成酶、转糖苷酶和糖基水解酶的作用下构建成兼具坚固性和柔韧性的细胞壁.三维网络状结构的细胞壁一直处于动态变化中,其组分随着形态发生而发生改变,如休眠期的孢子具有黑色素和rodlet输水蛋白层,在营养菌丝生长过程中这些被逐渐降解,同时出现菌丝特有组分半乳糖氨基半乳聚糖(GAG)(Fontaine et al. 2011;Briard et al. 2016)(图1). ...
... 另一种含半乳糖的聚合物是半乳糖氨基半乳聚糖(GAG),是只在营养菌丝体中发现的表面多糖.这种糖聚合物呈线性不溶于水,平均大小为100kDa,由半乳吡喃糖,半乳糖胺和N-乙酰半乳糖胺(GalNAc)以α-1,4 -糖苷键连接而成,并沿着多糖链随机分布(Fontaine et al. 2011).与其他糖聚合物一样,GAG的合成使用UDP-半乳糖和UDP-GalNAc为底物(Briard et al. 2016),并推测出几个与GAG合成相关的酶(Bamford et al. 2015;Gravelat et al. 2013).最近蛋白质拓扑结构预测显示CAZy家族 (http://www.cazy.org)GT4 的一种α-糖基转移酶有14个跨质膜结构域,可能是GAG合成酶(Briard et al. 2016).然而目前尚没有GAG合成酶的体外活性证明,这种酶是如何识别并连接GAG中不同的糖仍然是个谜. ...
... ).最近蛋白质拓扑结构预测显示CAZy家族 (http://www.cazy.org)GT4 的一种α-糖基转移酶有14个跨质膜结构域,可能是GAG合成酶(Briard et al. 2016).然而目前尚没有GAG合成酶的体外活性证明,这种酶是如何识别并连接GAG中不同的糖仍然是个谜. ...
Crh1p and Crh2p are required for the cross-linking of chitin to beta(1-6)glucan in the Saccharomyces cerevisiae cell wall.
1
2007
... 细胞壁多糖到达细胞壁空间后的最后一步是各个组分之间的交联.行驶交联功能的酶通常是通过GPI锚定在细胞壁上的蛋白.酵母中发现CRH家族负责几丁质与葡聚糖之间的交联,CRH缺失后的突变株表现出子细胞与母细胞连接处形态的异常(Cabib et al. 2007;Blanco et al. 2012).烟曲霉CRH家族有5个成员,我们最近的实验结果表明:整个家族的五重突变株未见任何菌丝体或分生孢子表型(未发表文章),让人开始质疑 β-葡聚糖-几丁质交联在烟曲霉细胞壁结构化中的作用. ...
Deciphering cell wall integrity signalling in Aspergillus fumigatus: identification and functional characterization of cell wall stress sensors and relevant Rho GTPases.
1
2012
... β-1,3-葡聚糖的合成以细胞内葡萄糖的活化形式UDP-Glc为合成前体,由跨膜葡聚糖合成酶复合体催化合成.早在1993年Latgé实验室就利用放射性标记的UDP(14C)-glucose从烟曲霉菌丝体膜组分中提取到了葡聚糖合成酶复合体,发现其由催化亚基和调控亚基组成(Beauvais et al. 2001).催化亚基为具有16个跨膜结构域的膜蛋白,是棘白菌素类药物的靶标(Douglas et al. 1997).催化亚基由基因fks1编码,在烟曲霉基因组中仅有一个拷贝.最早的研究表明 fks1是重要基因(Firon et al. 2002;Mouyna et al. 2004),然而Dichtl et al.(2015)最近发现fks1不是必需的,但其缺失突变株的菌落生长非常缓慢,表现为细胞壁β-1,3-葡聚糖缺失伴随着几丁质和半乳糖氨基半乳聚糖的补偿性增加以及甘露聚糖含量的显著下降.这可能是棘白菌素类药物只能抑制但不能杀死烟曲霉的原因.烟曲霉Rho-GTPase Rho1被鉴定为FKS1的调控亚基(Beauvais et al. 2001),遗传学实验表明它也是必需基因,缺失后导致细胞壁完整性途径(CWI)受损,细胞质渗漏裂解及最终不能存活(Dichtl et al. 2012). ...
Aspergillus fumigatus devoid of cell wall beta-1,3-glucan is viable, massively sheds galactomannan and is killed by septum formation inhibitors
1
2015
... β-1,3-葡聚糖的合成以细胞内葡萄糖的活化形式UDP-Glc为合成前体,由跨膜葡聚糖合成酶复合体催化合成.早在1993年Latgé实验室就利用放射性标记的UDP(14C)-glucose从烟曲霉菌丝体膜组分中提取到了葡聚糖合成酶复合体,发现其由催化亚基和调控亚基组成(Beauvais et al. 2001).催化亚基为具有16个跨膜结构域的膜蛋白,是棘白菌素类药物的靶标(Douglas et al. 1997).催化亚基由基因fks1编码,在烟曲霉基因组中仅有一个拷贝.最早的研究表明 fks1是重要基因(Firon et al. 2002;Mouyna et al. 2004),然而Dichtl et al.(2015)最近发现fks1不是必需的,但其缺失突变株的菌落生长非常缓慢,表现为细胞壁β-1,3-葡聚糖缺失伴随着几丁质和半乳糖氨基半乳聚糖的补偿性增加以及甘露聚糖含量的显著下降.这可能是棘白菌素类药物只能抑制但不能杀死烟曲霉的原因.烟曲霉Rho-GTPase Rho1被鉴定为FKS1的调控亚基(Beauvais et al. 2001),遗传学实验表明它也是必需基因,缺失后导致细胞壁完整性途径(CWI)受损,细胞质渗漏裂解及最终不能存活(Dichtl et al. 2012). ...
Identification of the FKS1 gene of Candida albicans as the essential target of 1,3-beta-D-glucan synthase inhibitors.
1
1997
... β-1,3-葡聚糖的合成以细胞内葡萄糖的活化形式UDP-Glc为合成前体,由跨膜葡聚糖合成酶复合体催化合成.早在1993年Latgé实验室就利用放射性标记的UDP(14C)-glucose从烟曲霉菌丝体膜组分中提取到了葡聚糖合成酶复合体,发现其由催化亚基和调控亚基组成(Beauvais et al. 2001).催化亚基为具有16个跨膜结构域的膜蛋白,是棘白菌素类药物的靶标(Douglas et al. 1997).催化亚基由基因fks1编码,在烟曲霉基因组中仅有一个拷贝.最早的研究表明 fks1是重要基因(Firon et al. 2002;Mouyna et al. 2004),然而Dichtl et al.(2015)最近发现fks1不是必需的,但其缺失突变株的菌落生长非常缓慢,表现为细胞壁β-1,3-葡聚糖缺失伴随着几丁质和半乳糖氨基半乳聚糖的补偿性增加以及甘露聚糖含量的显著下降.这可能是棘白菌素类药物只能抑制但不能杀死烟曲霉的原因.烟曲霉Rho-GTPase Rho1被鉴定为FKS1的调控亚基(Beauvais et al. 2001),遗传学实验表明它也是必需基因,缺失后导致细胞壁完整性途径(CWI)受损,细胞质渗漏裂解及最终不能存活(Dichtl et al. 2012). ...
Biosynthesis of the fungal cell wall polysaccharide galactomannan requires intraluminal GDP-mannose
1
2012
... 半乳甘露聚糖的合成需要甘露糖基转移酶和半乳糖基转移酶的协同作用,组装线性的α-甘露聚糖骨架和β-1,5-连接的半乳呋喃糖残基短链.烟曲霉的甘露聚糖合成以GDP-甘露糖为底物,由甘露糖基转移酶催化合成.每个甘露聚糖骨架平均有50个甘露糖残基,由多个α-连接的-M2-M2-M6- M2-重复单元组成(Latge et al. 1994),与葡聚糖-几丁质核心骨架共价连接.目前发现烟曲霉有11个甘露糖基转移酶负责α-1,6-和α-1,2-甘露糖苷键连接,但完全敲除这11个酶并没有导致菌丝体细胞壁中甘露聚糖含量的降低,仅分生孢子细胞壁的甘露聚糖含量减少(Henry et al. 2016).烟曲霉半乳甘露聚糖的半乳糖以半乳呋喃糖(Glaf)的形式存在,这种糖在许多致病菌和寄生虫中是很重要的己糖构型方式(Latge et al. 1994).UDP-Galf是半乳甘露聚糖的合成前体(Lamarre et al. 2009),由UDP-半乳吡喃糖在UDP-半乳糖变位酶的催化下转化而成.半乳甘露聚糖的合成最初是在内质网中进行,随后在高尔基复合体中进一步延伸并共价结合β-1,3-葡聚糖(Engel et al. 2012). ...
Identification of Aspergillus fumigatus cell wall as an anti-fungal target.
1
2017
... 法国巴斯德研究所曲霉中心Jean-Paul Latgé实验室30多年来一直致力于烟曲霉细胞壁的研究.经过多年来的努力目前确定烟曲霉细胞壁主要组分为碱不溶性的纤维状骨架,包括几丁质、甘露聚糖、β-1,3-葡聚糖以及β-1,6-分支的β-1,3-葡聚糖,包埋于无定形的碱可溶组分中包括α-1,3-葡聚糖和半乳甘露聚糖(Fontaine et al. 2000).烟曲霉细胞壁约占细胞干重的30%,各多糖组分含量大约为7%-15%的几丁质、20%-35%的β-1,3-葡聚糖、35%-46%的α-1,3-葡聚糖和20%-25%的半乳甘露聚糖(Gastebois et al. 2009).多糖的生物合成起始于胞内活化的核苷酸糖,参与核苷酸糖合成的酶为潜在的药物靶标(Fang et al. 2013a,2013b;Fang 2017).这些核苷酸糖在一系列跨膜合成酶、转糖苷酶和糖基水解酶的作用下构建成兼具坚固性和柔韧性的细胞壁.三维网络状结构的细胞壁一直处于动态变化中,其组分随着形态发生而发生改变,如休眠期的孢子具有黑色素和rodlet输水蛋白层,在营养菌丝生长过程中这些被逐渐降解,同时出现菌丝特有组分半乳糖氨基半乳聚糖(GAG)(Fontaine et al. 2011;Briard et al. 2016)(图1). ...
Genetic and structural validation of Aspergillus fumigatus N-acetylphosphoglucosamine mutase as an antifungal target.
1
2013
... 法国巴斯德研究所曲霉中心Jean-Paul Latgé实验室30多年来一直致力于烟曲霉细胞壁的研究.经过多年来的努力目前确定烟曲霉细胞壁主要组分为碱不溶性的纤维状骨架,包括几丁质、甘露聚糖、β-1,3-葡聚糖以及β-1,6-分支的β-1,3-葡聚糖,包埋于无定形的碱可溶组分中包括α-1,3-葡聚糖和半乳甘露聚糖(Fontaine et al. 2000).烟曲霉细胞壁约占细胞干重的30%,各多糖组分含量大约为7%-15%的几丁质、20%-35%的β-1,3-葡聚糖、35%-46%的α-1,3-葡聚糖和20%-25%的半乳甘露聚糖(Gastebois et al. 2009).多糖的生物合成起始于胞内活化的核苷酸糖,参与核苷酸糖合成的酶为潜在的药物靶标(Fang et al. 2013a,2013b;Fang 2017).这些核苷酸糖在一系列跨膜合成酶、转糖苷酶和糖基水解酶的作用下构建成兼具坚固性和柔韧性的细胞壁.三维网络状结构的细胞壁一直处于动态变化中,其组分随着形态发生而发生改变,如休眠期的孢子具有黑色素和rodlet输水蛋白层,在营养菌丝生长过程中这些被逐渐降解,同时出现菌丝特有组分半乳糖氨基半乳聚糖(GAG)(Fontaine et al. 2011;Briard et al. 2016)(图1). ...
Genetic and structural validation of Aspergillus fumigatus UDP-N-acetylglucosamine pyrophosphorylase as an antifungal target.
1
2013
... 法国巴斯德研究所曲霉中心Jean-Paul Latgé实验室30多年来一直致力于烟曲霉细胞壁的研究.经过多年来的努力目前确定烟曲霉细胞壁主要组分为碱不溶性的纤维状骨架,包括几丁质、甘露聚糖、β-1,3-葡聚糖以及β-1,6-分支的β-1,3-葡聚糖,包埋于无定形的碱可溶组分中包括α-1,3-葡聚糖和半乳甘露聚糖(Fontaine et al. 2000).烟曲霉细胞壁约占细胞干重的30%,各多糖组分含量大约为7%-15%的几丁质、20%-35%的β-1,3-葡聚糖、35%-46%的α-1,3-葡聚糖和20%-25%的半乳甘露聚糖(Gastebois et al. 2009).多糖的生物合成起始于胞内活化的核苷酸糖,参与核苷酸糖合成的酶为潜在的药物靶标(Fang et al. 2013a,2013b;Fang 2017).这些核苷酸糖在一系列跨膜合成酶、转糖苷酶和糖基水解酶的作用下构建成兼具坚固性和柔韧性的细胞壁.三维网络状结构的细胞壁一直处于动态变化中,其组分随着形态发生而发生改变,如休眠期的孢子具有黑色素和rodlet输水蛋白层,在营养菌丝生长过程中这些被逐渐降解,同时出现菌丝特有组分半乳糖氨基半乳聚糖(GAG)(Fontaine et al. 2011;Briard et al. 2016)(图1). ...
Characterization of essential genes by parasexual genetics in the human fungal pathogen Aspergillus fumigatus: impact of genomic rearrangements associated with electroporation of DNA.
1
2002
... β-1,3-葡聚糖的合成以细胞内葡萄糖的活化形式UDP-Glc为合成前体,由跨膜葡聚糖合成酶复合体催化合成.早在1993年Latgé实验室就利用放射性标记的UDP(14C)-glucose从烟曲霉菌丝体膜组分中提取到了葡聚糖合成酶复合体,发现其由催化亚基和调控亚基组成(Beauvais et al. 2001).催化亚基为具有16个跨膜结构域的膜蛋白,是棘白菌素类药物的靶标(Douglas et al. 1997).催化亚基由基因fks1编码,在烟曲霉基因组中仅有一个拷贝.最早的研究表明 fks1是重要基因(Firon et al. 2002;Mouyna et al. 2004),然而Dichtl et al.(2015)最近发现fks1不是必需的,但其缺失突变株的菌落生长非常缓慢,表现为细胞壁β-1,3-葡聚糖缺失伴随着几丁质和半乳糖氨基半乳聚糖的补偿性增加以及甘露聚糖含量的显著下降.这可能是棘白菌素类药物只能抑制但不能杀死烟曲霉的原因.烟曲霉Rho-GTPase Rho1被鉴定为FKS1的调控亚基(Beauvais et al. 2001),遗传学实验表明它也是必需基因,缺失后导致细胞壁完整性途径(CWI)受损,细胞质渗漏裂解及最终不能存活(Dichtl et al. 2012). ...
Galactosaminogalactan, a new immunosuppressive polysaccharide of Aspergillus fumigatus.
3
2011
... 法国巴斯德研究所曲霉中心Jean-Paul Latgé实验室30多年来一直致力于烟曲霉细胞壁的研究.经过多年来的努力目前确定烟曲霉细胞壁主要组分为碱不溶性的纤维状骨架,包括几丁质、甘露聚糖、β-1,3-葡聚糖以及β-1,6-分支的β-1,3-葡聚糖,包埋于无定形的碱可溶组分中包括α-1,3-葡聚糖和半乳甘露聚糖(Fontaine et al. 2000).烟曲霉细胞壁约占细胞干重的30%,各多糖组分含量大约为7%-15%的几丁质、20%-35%的β-1,3-葡聚糖、35%-46%的α-1,3-葡聚糖和20%-25%的半乳甘露聚糖(Gastebois et al. 2009).多糖的生物合成起始于胞内活化的核苷酸糖,参与核苷酸糖合成的酶为潜在的药物靶标(Fang et al. 2013a,2013b;Fang 2017).这些核苷酸糖在一系列跨膜合成酶、转糖苷酶和糖基水解酶的作用下构建成兼具坚固性和柔韧性的细胞壁.三维网络状结构的细胞壁一直处于动态变化中,其组分随着形态发生而发生改变,如休眠期的孢子具有黑色素和rodlet输水蛋白层,在营养菌丝生长过程中这些被逐渐降解,同时出现菌丝特有组分半乳糖氨基半乳聚糖(GAG)(Fontaine et al. 2011;Briard et al. 2016)(图1). ...
... 另一种含半乳糖的聚合物是半乳糖氨基半乳聚糖(GAG),是只在营养菌丝体中发现的表面多糖.这种糖聚合物呈线性不溶于水,平均大小为100kDa,由半乳吡喃糖,半乳糖胺和N-乙酰半乳糖胺(GalNAc)以α-1,4 -糖苷键连接而成,并沿着多糖链随机分布(Fontaine et al. 2011).与其他糖聚合物一样,GAG的合成使用UDP-半乳糖和UDP-GalNAc为底物(Briard et al. 2016),并推测出几个与GAG合成相关的酶(Bamford et al. 2015;Gravelat et al. 2013).最近蛋白质拓扑结构预测显示CAZy家族 (http://www.cazy.org)GT4 的一种α-糖基转移酶有14个跨质膜结构域,可能是GAG合成酶(Briard et al. 2016).然而目前尚没有GAG合成酶的体外活性证明,这种酶是如何识别并连接GAG中不同的糖仍然是个谜. ...
... GAG是菌丝特有的组分,参与生物膜的形成(Fontaine et al. 2011),能促进菌丝与巨噬细胞、嗜中性粒细胞和血小板的结合(Rambach et al. 2015).另外还具有以下的直接免疫抑制作用:(1)通过NK细胞依赖性机制引发多形核中性粒细胞(polymorphonuclear neutrophil,PMN)凋亡并抑制PMN趋化性(Robinet et al. 2014);(2)介导对依赖于NADPH氧化酶的嗜中性粒细胞细胞外捕获的抗性(Lee et al. 2015).GM在菌丝生长过程中被分泌出来,是用于IA早期诊断的有效抗原之一(Foy et al. 2007).Bio-Rad开发的基于GM单克隆抗体的PlateliaTM检测试剂盒因其较好的特异性和灵敏性被广泛用于临床检测中.最近有报道称,Dectin-2与GM结合(Reedy et al. 2016),但这种C型凝集素的免疫功能尚不完全清楚. ...
Molecular organization of the alkali-insoluble fraction of Aspergillus fumigatus cell wall.
3
2000
... 传统观念认为细胞壁是一种细胞外惰性骨架结构,然而近年来对机会致病真菌烟曲霉的细胞壁研究表明:细胞壁是个活的动态细胞器.烟曲霉是一种普遍存在于环境中的腐生丝状真菌,同时也是三大机会致病真菌之一,在免疫系统低下或缺陷的人群中可引起多种急慢性疾病.烟曲霉有很强的环境适应性,其细胞壁组分随着细胞周期进程和外部环境的变化而变化,这对理解细胞壁的结构组成和分子合成机制提出了重大挑战.研究表明,烟曲霉细胞壁主要由糖蛋白和多糖组成,其中多糖含量高达90%(Fontaine et al. 2000).鉴定细胞壁各组分使用方法是碱处理细胞壁后,将碱可溶与碱不溶组分分离,再用酶解或酸水解的方法处理两种组分,最后用经典的糖分析鉴定方法测定单糖或多糖的含量,最终确定细胞壁的组分及组成(Fontaine et al. 2000). ...
... ).鉴定细胞壁各组分使用方法是碱处理细胞壁后,将碱可溶与碱不溶组分分离,再用酶解或酸水解的方法处理两种组分,最后用经典的糖分析鉴定方法测定单糖或多糖的含量,最终确定细胞壁的组分及组成(Fontaine et al. 2000). ...
... 法国巴斯德研究所曲霉中心Jean-Paul Latgé实验室30多年来一直致力于烟曲霉细胞壁的研究.经过多年来的努力目前确定烟曲霉细胞壁主要组分为碱不溶性的纤维状骨架,包括几丁质、甘露聚糖、β-1,3-葡聚糖以及β-1,6-分支的β-1,3-葡聚糖,包埋于无定形的碱可溶组分中包括α-1,3-葡聚糖和半乳甘露聚糖(Fontaine et al. 2000).烟曲霉细胞壁约占细胞干重的30%,各多糖组分含量大约为7%-15%的几丁质、20%-35%的β-1,3-葡聚糖、35%-46%的α-1,3-葡聚糖和20%-25%的半乳甘露聚糖(Gastebois et al. 2009).多糖的生物合成起始于胞内活化的核苷酸糖,参与核苷酸糖合成的酶为潜在的药物靶标(Fang et al. 2013a,2013b;Fang 2017).这些核苷酸糖在一系列跨膜合成酶、转糖苷酶和糖基水解酶的作用下构建成兼具坚固性和柔韧性的细胞壁.三维网络状结构的细胞壁一直处于动态变化中,其组分随着形态发生而发生改变,如休眠期的孢子具有黑色素和rodlet输水蛋白层,在营养菌丝生长过程中这些被逐渐降解,同时出现菌丝特有组分半乳糖氨基半乳聚糖(GAG)(Fontaine et al. 2011;Briard et al. 2016)(图1). ...
Galactomannan antigen enzyme-linked immunosorbent assay for diagnosis of invasive aspergillosis after hematopoietic stem cell transplantation
0
2007
Evidence that the Aspergillus nidulans class I and class II chitin synthase genes, chsC and chsA, share critical roles in hyphal wall integrity and conidiophore development.
1
2000
... 几丁质是β-1,4-连接的N-乙酰葡萄糖胺线性聚合而成.已知几丁质由跨膜几丁质合成酶(Chitin synthase,CHS)以UDP- N-乙酰葡萄糖胺为前体合成.但几丁质合成机理尚不清楚,原因之一是丝状真菌中 CHS基因数量较多(Muszkieta et al. 2014).基于生物信息学分析烟曲霉有8个chs基因分属于两个家族.家族一的chsA、chsB和chsC对几丁质的合成贡献不大,但chsG单突变株及与chsA/B/C的多重突变株有分生孢子减少,菌丝形态改变、生长下降和几丁质合成酶活性降低等表型(Mellado et al. 1996;Mellado et al. 2003).家族二包括csmA、csmB、chsF和chsD 4个基因,4重敲除株表现出菌丝形态和孢子的变化,但几丁质含量几乎不受影响.敲除两个家族的4个基因(ΔcsmA/ΔcsmB/ ΔchsF/ΔchsG)后菌丝生长非常缓慢(Muszkieta et al. 2014),类似于fks1突变株,表明两个家族的几丁质合成酶共同协作参与烟曲霉几丁质的合成,也表明几丁质的合成对菌丝生长的重要性.构巢曲霉的细胞壁结构和烟曲霉的非常相似,但其几丁质合成酶的分类和功能与烟曲霉不同(Fujiwara et al. 2000;Tsuizaki et al. 2009),说明不同真菌中几丁质合成酶的功能根据细胞壁整体合成调控的需求有所不同. ...
Aspergillus fumigatus: cell wall polysaccharides, their biosynthesis and organization
1
2009
... 法国巴斯德研究所曲霉中心Jean-Paul Latgé实验室30多年来一直致力于烟曲霉细胞壁的研究.经过多年来的努力目前确定烟曲霉细胞壁主要组分为碱不溶性的纤维状骨架,包括几丁质、甘露聚糖、β-1,3-葡聚糖以及β-1,6-分支的β-1,3-葡聚糖,包埋于无定形的碱可溶组分中包括α-1,3-葡聚糖和半乳甘露聚糖(Fontaine et al. 2000).烟曲霉细胞壁约占细胞干重的30%,各多糖组分含量大约为7%-15%的几丁质、20%-35%的β-1,3-葡聚糖、35%-46%的α-1,3-葡聚糖和20%-25%的半乳甘露聚糖(Gastebois et al. 2009).多糖的生物合成起始于胞内活化的核苷酸糖,参与核苷酸糖合成的酶为潜在的药物靶标(Fang et al. 2013a,2013b;Fang 2017).这些核苷酸糖在一系列跨膜合成酶、转糖苷酶和糖基水解酶的作用下构建成兼具坚固性和柔韧性的细胞壁.三维网络状结构的细胞壁一直处于动态变化中,其组分随着形态发生而发生改变,如休眠期的孢子具有黑色素和rodlet输水蛋白层,在营养菌丝生长过程中这些被逐渐降解,同时出现菌丝特有组分半乳糖氨基半乳聚糖(GAG)(Fontaine et al. 2011;Briard et al. 2016)(图1). ...
beta(1-3)glucanosyltransferase Gel4p is essential for Aspergillus fumigatus.
1
2010
... 新合成的多糖链在细胞壁空间需要特定糖基转移酶和水解酶的共同作用进行重构、交联以产生细胞壁的三维网络结构,其中以β-1,3-葡聚糖的修饰最为多样.由葡聚糖合成酶复合体合成的线性β-1,3-葡聚糖需要经过延伸、分支和水解等过程才构成细胞骨架.葡聚糖的延伸是由糖基转移酶GEL家族负责,将水解后糖链的还原端转运到另一个葡聚糖分子的非还原端,从而延伸葡聚糖链.GEL属于CAZy数据库中的GH72家族,在烟曲霉中有7个成员,其中GEL4的缺失是致死的(Mouyna et al. 2005;Gastebois et al. 2010a),其他成员对葡聚糖的合成、形态发生、细胞壁完整性途径和致病力有不同程度的影响(Mouyna et al. 2005;Zhao et al. 2014).最近的研究表明含有碳水化合物结合域(carbohydrate binding module,CBM)的GEL4除了延伸β-1,3-葡聚糖外还可以建立分支,具有双重功能(Aimanianda et al. 2017).BGT1和BGT2基因也具有β-1,3-葡聚糖的分支功能,可以产生β-1,6-分支的β-1,3-葡聚糖,但它们的缺失没有引起细胞壁表型(Gastebois et al. 2010b),说明在细胞壁组装中它们并非重要基因.孢子萌发或菌丝分支生长中需要葡萄糖苷酶水解β-1,3-葡聚糖以增加细胞壁的可塑性.烟曲霉基因组含有多个β-1,3-葡聚糖酶,目前对其中eng1和eng2基因的功能分析发现它们的缺失未导致任何细胞壁缺陷(Mouyna et al. 2002;Hartl et al. 2011). ...
Characterization of a new beta(1-3)-glucan branching activity of Aspergillus fumigatus.
1
2010
... 新合成的多糖链在细胞壁空间需要特定糖基转移酶和水解酶的共同作用进行重构、交联以产生细胞壁的三维网络结构,其中以β-1,3-葡聚糖的修饰最为多样.由葡聚糖合成酶复合体合成的线性β-1,3-葡聚糖需要经过延伸、分支和水解等过程才构成细胞骨架.葡聚糖的延伸是由糖基转移酶GEL家族负责,将水解后糖链的还原端转运到另一个葡聚糖分子的非还原端,从而延伸葡聚糖链.GEL属于CAZy数据库中的GH72家族,在烟曲霉中有7个成员,其中GEL4的缺失是致死的(Mouyna et al. 2005;Gastebois et al. 2010a),其他成员对葡聚糖的合成、形态发生、细胞壁完整性途径和致病力有不同程度的影响(Mouyna et al. 2005;Zhao et al. 2014).最近的研究表明含有碳水化合物结合域(carbohydrate binding module,CBM)的GEL4除了延伸β-1,3-葡聚糖外还可以建立分支,具有双重功能(Aimanianda et al. 2017).BGT1和BGT2基因也具有β-1,3-葡聚糖的分支功能,可以产生β-1,6-分支的β-1,3-葡聚糖,但它们的缺失没有引起细胞壁表型(Gastebois et al. 2010b),说明在细胞壁组装中它们并非重要基因.孢子萌发或菌丝分支生长中需要葡萄糖苷酶水解β-1,3-葡聚糖以增加细胞壁的可塑性.烟曲霉基因组含有多个β-1,3-葡聚糖酶,目前对其中eng1和eng2基因的功能分析发现它们的缺失未导致任何细胞壁缺陷(Mouyna et al. 2002;Hartl et al. 2011). ...
Dectin-1 and TLRs permit macrophages to distinguish between different Aspergillus fumigatus cellular states.
1
2006
... 细胞壁含有病原体相关分子模式(pathogen- associated molecular patterns,PAMPs),可被宿主先天免疫细胞的模式识别受体(pattern recognition receptors,PRR)识别,这对免疫反应的启动是必不可少的.目前发现的PRR包括Toll样受体(TLRs),C-型凝集素受体(CLRs),NOD样受体(NLRs)和可溶性PRRs.研究结果表明:细胞壁的β-1,3-葡聚糖可以被CLRs中的Dectin-1特异性识别(Herre et al. 2004),从而激活了Dectin-1信号通路(Gersuk et al. 2006)和PRRs调节吞噬体成熟的能力,初步揭示了真菌利用PAMPs逃离巨噬细胞杀伤的机制(Mansour et al. 2013). ...
Aspergillus galactosaminogalactan mediates adherence to host constituents and conceals hyphal beta-glucan from the immune system
2
2013
... 另一种含半乳糖的聚合物是半乳糖氨基半乳聚糖(GAG),是只在营养菌丝体中发现的表面多糖.这种糖聚合物呈线性不溶于水,平均大小为100kDa,由半乳吡喃糖,半乳糖胺和N-乙酰半乳糖胺(GalNAc)以α-1,4 -糖苷键连接而成,并沿着多糖链随机分布(Fontaine et al. 2011).与其他糖聚合物一样,GAG的合成使用UDP-半乳糖和UDP-GalNAc为底物(Briard et al. 2016),并推测出几个与GAG合成相关的酶(Bamford et al. 2015;Gravelat et al. 2013).最近蛋白质拓扑结构预测显示CAZy家族 (http://www.cazy.org)GT4 的一种α-糖基转移酶有14个跨质膜结构域,可能是GAG合成酶(Briard et al. 2016).然而目前尚没有GAG合成酶的体外活性证明,这种酶是如何识别并连接GAG中不同的糖仍然是个谜. ...
... 分生孢子在萌发时首先是经过膨胀阶段,此时rodlet层被天冬氨酸蛋白酶降解,疏水性孢子开始变成亲水性的.随着细胞内渗透压的增加孢子开始膨大,细胞壁多糖被糖基水解酶降解并伴有新的多糖层开始合成.膨胀的孢子转向极性生长时芽管开始出现,此时黑色素层被瓦解,膨胀期合成的多糖层构成了菌丝的细胞壁,随后有细胞外基质成分GAG覆盖于新的细胞壁表面(Gravelat et al. 2013). ...
Characterization of the GPI-anchored endo beta-1,3-glucanase Eng2 of Aspergillus fumigatus.
1
2011
... 新合成的多糖链在细胞壁空间需要特定糖基转移酶和水解酶的共同作用进行重构、交联以产生细胞壁的三维网络结构,其中以β-1,3-葡聚糖的修饰最为多样.由葡聚糖合成酶复合体合成的线性β-1,3-葡聚糖需要经过延伸、分支和水解等过程才构成细胞骨架.葡聚糖的延伸是由糖基转移酶GEL家族负责,将水解后糖链的还原端转运到另一个葡聚糖分子的非还原端,从而延伸葡聚糖链.GEL属于CAZy数据库中的GH72家族,在烟曲霉中有7个成员,其中GEL4的缺失是致死的(Mouyna et al. 2005;Gastebois et al. 2010a),其他成员对葡聚糖的合成、形态发生、细胞壁完整性途径和致病力有不同程度的影响(Mouyna et al. 2005;Zhao et al. 2014).最近的研究表明含有碳水化合物结合域(carbohydrate binding module,CBM)的GEL4除了延伸β-1,3-葡聚糖外还可以建立分支,具有双重功能(Aimanianda et al. 2017).BGT1和BGT2基因也具有β-1,3-葡聚糖的分支功能,可以产生β-1,6-分支的β-1,3-葡聚糖,但它们的缺失没有引起细胞壁表型(Gastebois et al. 2010b),说明在细胞壁组装中它们并非重要基因.孢子萌发或菌丝分支生长中需要葡萄糖苷酶水解β-1,3-葡聚糖以增加细胞壁的可塑性.烟曲霉基因组含有多个β-1,3-葡聚糖酶,目前对其中eng1和eng2基因的功能分析发现它们的缺失未导致任何细胞壁缺陷(Mouyna et al. 2002;Hartl et al. 2011). ...
Aspergillus fumigatus dihydroxynaphthalene melanin modulates the host endocytosis pathway
1
2012
... 烟曲霉有3种细胞形态:营养菌丝体形态,主要分解土壤中的有机物质维持碳氮循环;无性分生孢子形态,主要介导在空气中的传播;休眠的囊孢子状态,确保自身的长期生存.分生孢子和菌丝体的细胞壁结构组分非常相似,但两种形态又有各自特有的细胞壁外层组分(图1B、图1C)(Latge & Beauvais 2014).分生孢子的外层有黑色素层,且被rodlet层覆盖.这个rodlet层完全是由 roda基因编码的疏水蛋白构成,具有8个半胱氨酸残基形成的4个二硫键排列成淀粉样从而使分生孢子细胞壁疏水(Aimanianda et al. 2009).位于rodlet层下面的是二羟基萘(DHN)黑色素层.基因敲除实验表明黑色素决定了孢子细胞壁的结构和硬度(Heinekamp et al. 2012;Bayry et al. 2014).最近Latgé实验室构建的ags1-3、roda和pks五重缺失株产生白色亲水性分生孢子且数量急剧减少,说明孢子外层的这些组分与产孢能力密切相关.五重缺失株的菌丝体生长不受影响,表明碱不溶性多糖组分是菌丝体细胞壁必需的(Latge et al. 2017). ...
Biosynthesis of cell wall mannan in the conidium and the mycelium of Aspergillus fumigatus.
1
2016
... 半乳甘露聚糖的合成需要甘露糖基转移酶和半乳糖基转移酶的协同作用,组装线性的α-甘露聚糖骨架和β-1,5-连接的半乳呋喃糖残基短链.烟曲霉的甘露聚糖合成以GDP-甘露糖为底物,由甘露糖基转移酶催化合成.每个甘露聚糖骨架平均有50个甘露糖残基,由多个α-连接的-M2-M2-M6- M2-重复单元组成(Latge et al. 1994),与葡聚糖-几丁质核心骨架共价连接.目前发现烟曲霉有11个甘露糖基转移酶负责α-1,6-和α-1,2-甘露糖苷键连接,但完全敲除这11个酶并没有导致菌丝体细胞壁中甘露聚糖含量的降低,仅分生孢子细胞壁的甘露聚糖含量减少(Henry et al. 2016).烟曲霉半乳甘露聚糖的半乳糖以半乳呋喃糖(Glaf)的形式存在,这种糖在许多致病菌和寄生虫中是很重要的己糖构型方式(Latge et al. 1994).UDP-Galf是半乳甘露聚糖的合成前体(Lamarre et al. 2009),由UDP-半乳吡喃糖在UDP-半乳糖变位酶的催化下转化而成.半乳甘露聚糖的合成最初是在内质网中进行,随后在高尔基复合体中进一步延伸并共价结合β-1,3-葡聚糖(Engel et al. 2012). ...
alpha1,3 glucans are dispensable in Aspergillus fumigatus.
2
2012
... α-1,3-葡聚糖是细胞壁的无定形组分,约占细胞壁含量的40%.烟曲霉有3种α-1,3-葡聚糖合成酶(AGS1、AGS2和AGS3),都是高分子量的跨膜酶 (>200kDa),包括一个淀粉酶样的结构域和一个似糖原合成酶样的结构域.3个基因在菌丝生长期均为高表达但都不是必需基因(Beauvais et al. 2005;Maubon et al. 2006;Henry et al. 2012).三重缺失株不含α-1,3-葡聚糖但菌丝营养生长不受影响,菌丝的形态通过β-1,3-葡聚糖和几丁质含量的增加来维持(Henry et al. 2012).目前对α-1,3-葡聚糖合成酶的底物和体外合成活性尚不清楚. ...
... ).三重缺失株不含α-1,3-葡聚糖但菌丝营养生长不受影响,菌丝的形态通过β-1,3-葡聚糖和几丁质含量的增加来维持(Henry et al. 2012).目前对α-1,3-葡聚糖合成酶的底物和体外合成活性尚不清楚. ...
Dectin-1 and its role in the recognition of beta-glucans by macrophages
1
2004
... 细胞壁含有病原体相关分子模式(pathogen- associated molecular patterns,PAMPs),可被宿主先天免疫细胞的模式识别受体(pattern recognition receptors,PRR)识别,这对免疫反应的启动是必不可少的.目前发现的PRR包括Toll样受体(TLRs),C-型凝集素受体(CLRs),NOD样受体(NLRs)和可溶性PRRs.研究结果表明:细胞壁的β-1,3-葡聚糖可以被CLRs中的Dectin-1特异性识别(Herre et al. 2004),从而激活了Dectin-1信号通路(Gersuk et al. 2006)和PRRs调节吞噬体成熟的能力,初步揭示了真菌利用PAMPs逃离巨噬细胞杀伤的机制(Mansour et al. 2013). ...
Disruption of the gene encoding the ChiB1 chitinase of Aspergillus fumigatus and characterization of a recombinant gene product.
1
2003
... 几丁质作为提供坚固性的细胞壁组分在形态发生中一直处于动态变化.几丁质酶水解β-1,4-糖苷键从而分解几丁质.烟曲霉有5个植物型、12个细菌型几丁质酶(Taib et al. 2005).植物型几丁质酶的五重敲除株与细菌型中AfChiB1的缺失都没有明显的表型(Jaques et al. 2003;Alcazar-Fuoli et al. 2011),说明不同类型的几丁质酶功能可以互补.另外几丁质脱乙酰酶和α-1,3-葡聚糖水解酶等在烟曲霉中也有多个冗余基因,利用遗传学技术分析这些酶的功能尚在进行中. ...
Galactofuranose attenuates cellular adhesion of Aspergillus fumigatus.
1
2009
... 半乳甘露聚糖的合成需要甘露糖基转移酶和半乳糖基转移酶的协同作用,组装线性的α-甘露聚糖骨架和β-1,5-连接的半乳呋喃糖残基短链.烟曲霉的甘露聚糖合成以GDP-甘露糖为底物,由甘露糖基转移酶催化合成.每个甘露聚糖骨架平均有50个甘露糖残基,由多个α-连接的-M2-M2-M6- M2-重复单元组成(Latge et al. 1994),与葡聚糖-几丁质核心骨架共价连接.目前发现烟曲霉有11个甘露糖基转移酶负责α-1,6-和α-1,2-甘露糖苷键连接,但完全敲除这11个酶并没有导致菌丝体细胞壁中甘露聚糖含量的降低,仅分生孢子细胞壁的甘露聚糖含量减少(Henry et al. 2016).烟曲霉半乳甘露聚糖的半乳糖以半乳呋喃糖(Glaf)的形式存在,这种糖在许多致病菌和寄生虫中是很重要的己糖构型方式(Latge et al. 1994).UDP-Galf是半乳甘露聚糖的合成前体(Lamarre et al. 2009),由UDP-半乳吡喃糖在UDP-半乳糖变位酶的催化下转化而成.半乳甘露聚糖的合成最初是在内质网中进行,随后在高尔基复合体中进一步延伸并共价结合β-1,3-葡聚糖(Engel et al. 2012). ...
Functional duality of the cell wall
2
2014
... 烟曲霉有3种细胞形态:营养菌丝体形态,主要分解土壤中的有机物质维持碳氮循环;无性分生孢子形态,主要介导在空气中的传播;休眠的囊孢子状态,确保自身的长期生存.分生孢子和菌丝体的细胞壁结构组分非常相似,但两种形态又有各自特有的细胞壁外层组分(图1B、图1C)(Latge & Beauvais 2014).分生孢子的外层有黑色素层,且被rodlet层覆盖.这个rodlet层完全是由 roda基因编码的疏水蛋白构成,具有8个半胱氨酸残基形成的4个二硫键排列成淀粉样从而使分生孢子细胞壁疏水(Aimanianda et al. 2009).位于rodlet层下面的是二羟基萘(DHN)黑色素层.基因敲除实验表明黑色素决定了孢子细胞壁的结构和硬度(Heinekamp et al. 2012;Bayry et al. 2014).最近Latgé实验室构建的ags1-3、roda和pks五重缺失株产生白色亲水性分生孢子且数量急剧减少,说明孢子外层的这些组分与产孢能力密切相关.五重缺失株的菌丝体生长不受影响,表明碱不溶性多糖组分是菌丝体细胞壁必需的(Latge et al. 2017). ...
... 五重缺失株产生白色亲水性分生孢子且数量急剧减少,说明孢子外层的这些组分与产孢能力密切相关.五重缺失株的菌丝体生长不受影响,表明碱不溶性多糖组分是菌丝体细胞壁必需的(Latge et al. 2017). ...
Chemical and immunological characterization of the extracellular galactomannan of Aspergillus fumigatus.
2
1994
... 半乳甘露聚糖的合成需要甘露糖基转移酶和半乳糖基转移酶的协同作用,组装线性的α-甘露聚糖骨架和β-1,5-连接的半乳呋喃糖残基短链.烟曲霉的甘露聚糖合成以GDP-甘露糖为底物,由甘露糖基转移酶催化合成.每个甘露聚糖骨架平均有50个甘露糖残基,由多个α-连接的-M2-M2-M6- M2-重复单元组成(Latge et al. 1994),与葡聚糖-几丁质核心骨架共价连接.目前发现烟曲霉有11个甘露糖基转移酶负责α-1,6-和α-1,2-甘露糖苷键连接,但完全敲除这11个酶并没有导致菌丝体细胞壁中甘露聚糖含量的降低,仅分生孢子细胞壁的甘露聚糖含量减少(Henry et al. 2016).烟曲霉半乳甘露聚糖的半乳糖以半乳呋喃糖(Glaf)的形式存在,这种糖在许多致病菌和寄生虫中是很重要的己糖构型方式(Latge et al. 1994).UDP-Galf是半乳甘露聚糖的合成前体(Lamarre et al. 2009),由UDP-半乳吡喃糖在UDP-半乳糖变位酶的催化下转化而成.半乳甘露聚糖的合成最初是在内质网中进行,随后在高尔基复合体中进一步延伸并共价结合β-1,3-葡聚糖(Engel et al. 2012). ...
... ).烟曲霉半乳甘露聚糖的半乳糖以半乳呋喃糖(Glaf)的形式存在,这种糖在许多致病菌和寄生虫中是很重要的己糖构型方式(Latge et al. 1994).UDP-Galf是半乳甘露聚糖的合成前体(Lamarre et al. 2009),由UDP-半乳吡喃糖在UDP-半乳糖变位酶的催化下转化而成.半乳甘露聚糖的合成最初是在内质网中进行,随后在高尔基复合体中进一步延伸并共价结合β-1,3-葡聚糖(Engel et al. 2012). ...
The Fungal exopolysaccharide galactosaminogalactan mediates virulence by enhancing resistance to neutrophil extracellular traps
1
2015
... GAG是菌丝特有的组分,参与生物膜的形成(Fontaine et al. 2011),能促进菌丝与巨噬细胞、嗜中性粒细胞和血小板的结合(Rambach et al. 2015).另外还具有以下的直接免疫抑制作用:(1)通过NK细胞依赖性机制引发多形核中性粒细胞(polymorphonuclear neutrophil,PMN)凋亡并抑制PMN趋化性(Robinet et al. 2014);(2)介导对依赖于NADPH氧化酶的嗜中性粒细胞细胞外捕获的抗性(Lee et al. 2015).GM在菌丝生长过程中被分泌出来,是用于IA早期诊断的有效抗原之一(Foy et al. 2007).Bio-Rad开发的基于GM单克隆抗体的PlateliaTM检测试剂盒因其较好的特异性和灵敏性被广泛用于临床检测中.最近有报道称,Dectin-2与GM结合(Reedy et al. 2016),但这种C型凝集素的免疫功能尚不完全清楚. ...
Exploiting fungal cell wall components in vaccines
1
2015
... 几丁质长期以来在过敏和直接抗真菌响应方面发挥着重要作用.对几丁质的免疫应答是细胞类型特异性的,并随着几丁质颗粒的浓度和大小的不同而不同,在人和小鼠中有不一致的报道(Wagener et al. 2014).几丁质能诱导人几丁三糖糖苷酶产生,降解不溶性几丁质为可溶性壳寡糖,从而作为更强的免疫刺激剂(Wiesner et al. 2015)激活宿主免疫反应.在小鼠中,几丁质选择性诱导IL-10产生但不刺激促炎细胞因子(Wagener et al. 2014)的产生.不同宿主中免疫应答结果不一致的一个原因是目前没有几丁质与受体直接结合的证据.过去发现的几种几丁质受体(Levitz et al. 2015)诱导产生细胞因子的作用仍然存在争议. ...
Dectin-1 activation controls maturation of beta-1,3-glucan- containing phagosomes
1
2013
... 细胞壁含有病原体相关分子模式(pathogen- associated molecular patterns,PAMPs),可被宿主先天免疫细胞的模式识别受体(pattern recognition receptors,PRR)识别,这对免疫反应的启动是必不可少的.目前发现的PRR包括Toll样受体(TLRs),C-型凝集素受体(CLRs),NOD样受体(NLRs)和可溶性PRRs.研究结果表明:细胞壁的β-1,3-葡聚糖可以被CLRs中的Dectin-1特异性识别(Herre et al. 2004),从而激活了Dectin-1信号通路(Gersuk et al. 2006)和PRRs调节吞噬体成熟的能力,初步揭示了真菌利用PAMPs逃离巨噬细胞杀伤的机制(Mansour et al. 2013). ...
AGS3, an alpha(1-3)glucan synthase gene family member of Aspergillus fumigatus, modulates mycelium growth in the lung of experimentally infected mice.
1
2006
... α-1,3-葡聚糖是细胞壁的无定形组分,约占细胞壁含量的40%.烟曲霉有3种α-1,3-葡聚糖合成酶(AGS1、AGS2和AGS3),都是高分子量的跨膜酶 (>200kDa),包括一个淀粉酶样的结构域和一个似糖原合成酶样的结构域.3个基因在菌丝生长期均为高表达但都不是必需基因(Beauvais et al. 2005;Maubon et al. 2006;Henry et al. 2012).三重缺失株不含α-1,3-葡聚糖但菌丝营养生长不受影响,菌丝的形态通过β-1,3-葡聚糖和几丁质含量的增加来维持(Henry et al. 2012).目前对α-1,3-葡聚糖合成酶的底物和体外合成活性尚不清楚. ...
The Aspergillus fumigatus chsC and chsG genes encode class III chitin synthases with different functions.
1
1996
... 几丁质是β-1,4-连接的N-乙酰葡萄糖胺线性聚合而成.已知几丁质由跨膜几丁质合成酶(Chitin synthase,CHS)以UDP- N-乙酰葡萄糖胺为前体合成.但几丁质合成机理尚不清楚,原因之一是丝状真菌中 CHS基因数量较多(Muszkieta et al. 2014).基于生物信息学分析烟曲霉有8个chs基因分属于两个家族.家族一的chsA、chsB和chsC对几丁质的合成贡献不大,但chsG单突变株及与chsA/B/C的多重突变株有分生孢子减少,菌丝形态改变、生长下降和几丁质合成酶活性降低等表型(Mellado et al. 1996;Mellado et al. 2003).家族二包括csmA、csmB、chsF和chsD 4个基因,4重敲除株表现出菌丝形态和孢子的变化,但几丁质含量几乎不受影响.敲除两个家族的4个基因(ΔcsmA/ΔcsmB/ ΔchsF/ΔchsG)后菌丝生长非常缓慢(Muszkieta et al. 2014),类似于fks1突变株,表明两个家族的几丁质合成酶共同协作参与烟曲霉几丁质的合成,也表明几丁质的合成对菌丝生长的重要性.构巢曲霉的细胞壁结构和烟曲霉的非常相似,但其几丁质合成酶的分类和功能与烟曲霉不同(Fujiwara et al. 2000;Tsuizaki et al. 2009),说明不同真菌中几丁质合成酶的功能根据细胞壁整体合成调控的需求有所不同. ...
Cell wall biogenesis in a double chitin synthase mutant (chsG-/chsE-) of Aspergillus fumigatus.
1
2003
... 几丁质是β-1,4-连接的N-乙酰葡萄糖胺线性聚合而成.已知几丁质由跨膜几丁质合成酶(Chitin synthase,CHS)以UDP- N-乙酰葡萄糖胺为前体合成.但几丁质合成机理尚不清楚,原因之一是丝状真菌中 CHS基因数量较多(Muszkieta et al. 2014).基于生物信息学分析烟曲霉有8个chs基因分属于两个家族.家族一的chsA、chsB和chsC对几丁质的合成贡献不大,但chsG单突变株及与chsA/B/C的多重突变株有分生孢子减少,菌丝形态改变、生长下降和几丁质合成酶活性降低等表型(Mellado et al. 1996;Mellado et al. 2003).家族二包括csmA、csmB、chsF和chsD 4个基因,4重敲除株表现出菌丝形态和孢子的变化,但几丁质含量几乎不受影响.敲除两个家族的4个基因(ΔcsmA/ΔcsmB/ ΔchsF/ΔchsG)后菌丝生长非常缓慢(Muszkieta et al. 2014),类似于fks1突变株,表明两个家族的几丁质合成酶共同协作参与烟曲霉几丁质的合成,也表明几丁质的合成对菌丝生长的重要性.构巢曲霉的细胞壁结构和烟曲霉的非常相似,但其几丁质合成酶的分类和功能与烟曲霉不同(Fujiwara et al. 2000;Tsuizaki et al. 2009),说明不同真菌中几丁质合成酶的功能根据细胞壁整体合成调控的需求有所不同. ...
Essential role for neutrophils but not alveolar macrophages at early time points following Aspergillus fumigatus infection.
1
2009
... 细胞壁对真菌的生长和抵抗环境压力至关重要,同时它也是病原真菌与宿主免疫细胞最先接触的细胞器.先天性免疫反应对于有效及时地识别烟曲霉并诱导炎症网络清除非常重要(Becker et al. 2015).在免疫活性宿主中,肺泡巨噬细胞(AM)和嗜中性粒细胞每天需要清除数百至上千的烟曲霉分生孢子以防止肺部的炎性损伤(Underhill et al. 2015).肺泡巨噬细胞(alveolar macrophages,AM)是唯一可以进入肺泡腔的吞噬细胞而且是第一个接触吸入孢子的细胞.嗜中性粒细胞是烟曲霉的必需免疫效应细胞,因为严重嗜中性白血球减少症是侵袭性曲霉病发展的主要危险因素(Mircescu et al. 2009).微小的烟曲霉孢子被吸入宿主肺部时,细胞壁组分有的发挥激活宿主免疫反应的作用,有的则保护孢子并促进其在宿主中的定殖和生存(表1). ...
Gene silencing with RNA interference in the human pathogenic fungus Aspergillus fumigatus.
1
2004
... β-1,3-葡聚糖的合成以细胞内葡萄糖的活化形式UDP-Glc为合成前体,由跨膜葡聚糖合成酶复合体催化合成.早在1993年Latgé实验室就利用放射性标记的UDP(14C)-glucose从烟曲霉菌丝体膜组分中提取到了葡聚糖合成酶复合体,发现其由催化亚基和调控亚基组成(Beauvais et al. 2001).催化亚基为具有16个跨膜结构域的膜蛋白,是棘白菌素类药物的靶标(Douglas et al. 1997).催化亚基由基因fks1编码,在烟曲霉基因组中仅有一个拷贝.最早的研究表明 fks1是重要基因(Firon et al. 2002;Mouyna et al. 2004),然而Dichtl et al.(2015)最近发现fks1不是必需的,但其缺失突变株的菌落生长非常缓慢,表现为细胞壁β-1,3-葡聚糖缺失伴随着几丁质和半乳糖氨基半乳聚糖的补偿性增加以及甘露聚糖含量的显著下降.这可能是棘白菌素类药物只能抑制但不能杀死烟曲霉的原因.烟曲霉Rho-GTPase Rho1被鉴定为FKS1的调控亚基(Beauvais et al. 2001),遗传学实验表明它也是必需基因,缺失后导致细胞壁完整性途径(CWI)受损,细胞质渗漏裂解及最终不能存活(Dichtl et al. 2012). ...
Deletion of GEL2 encoding for a beta(1-3)glucanosyltransferase affects morphogenesis and virulence in Aspergillus fumigatus.
2
2005
... 新合成的多糖链在细胞壁空间需要特定糖基转移酶和水解酶的共同作用进行重构、交联以产生细胞壁的三维网络结构,其中以β-1,3-葡聚糖的修饰最为多样.由葡聚糖合成酶复合体合成的线性β-1,3-葡聚糖需要经过延伸、分支和水解等过程才构成细胞骨架.葡聚糖的延伸是由糖基转移酶GEL家族负责,将水解后糖链的还原端转运到另一个葡聚糖分子的非还原端,从而延伸葡聚糖链.GEL属于CAZy数据库中的GH72家族,在烟曲霉中有7个成员,其中GEL4的缺失是致死的(Mouyna et al. 2005;Gastebois et al. 2010a),其他成员对葡聚糖的合成、形态发生、细胞壁完整性途径和致病力有不同程度的影响(Mouyna et al. 2005;Zhao et al. 2014).最近的研究表明含有碳水化合物结合域(carbohydrate binding module,CBM)的GEL4除了延伸β-1,3-葡聚糖外还可以建立分支,具有双重功能(Aimanianda et al. 2017).BGT1和BGT2基因也具有β-1,3-葡聚糖的分支功能,可以产生β-1,6-分支的β-1,3-葡聚糖,但它们的缺失没有引起细胞壁表型(Gastebois et al. 2010b),说明在细胞壁组装中它们并非重要基因.孢子萌发或菌丝分支生长中需要葡萄糖苷酶水解β-1,3-葡聚糖以增加细胞壁的可塑性.烟曲霉基因组含有多个β-1,3-葡聚糖酶,目前对其中eng1和eng2基因的功能分析发现它们的缺失未导致任何细胞壁缺陷(Mouyna et al. 2002;Hartl et al. 2011). ...
... ),其他成员对葡聚糖的合成、形态发生、细胞壁完整性途径和致病力有不同程度的影响(Mouyna et al. 2005;Zhao et al. 2014).最近的研究表明含有碳水化合物结合域(carbohydrate binding module,CBM)的GEL4除了延伸β-1,3-葡聚糖外还可以建立分支,具有双重功能(Aimanianda et al. 2017).BGT1和BGT2基因也具有β-1,3-葡聚糖的分支功能,可以产生β-1,6-分支的β-1,3-葡聚糖,但它们的缺失没有引起细胞壁表型(Gastebois et al. 2010b),说明在细胞壁组装中它们并非重要基因.孢子萌发或菌丝分支生长中需要葡萄糖苷酶水解β-1,3-葡聚糖以增加细胞壁的可塑性.烟曲霉基因组含有多个β-1,3-葡聚糖酶,目前对其中eng1和eng2基因的功能分析发现它们的缺失未导致任何细胞壁缺陷(Mouyna et al. 2002;Hartl et al. 2011). ...
Molecular characterization of a cell wall-associated beta(1-3)endoglucanase of Aspergillus fumigatus.
1
2002
... 新合成的多糖链在细胞壁空间需要特定糖基转移酶和水解酶的共同作用进行重构、交联以产生细胞壁的三维网络结构,其中以β-1,3-葡聚糖的修饰最为多样.由葡聚糖合成酶复合体合成的线性β-1,3-葡聚糖需要经过延伸、分支和水解等过程才构成细胞骨架.葡聚糖的延伸是由糖基转移酶GEL家族负责,将水解后糖链的还原端转运到另一个葡聚糖分子的非还原端,从而延伸葡聚糖链.GEL属于CAZy数据库中的GH72家族,在烟曲霉中有7个成员,其中GEL4的缺失是致死的(Mouyna et al. 2005;Gastebois et al. 2010a),其他成员对葡聚糖的合成、形态发生、细胞壁完整性途径和致病力有不同程度的影响(Mouyna et al. 2005;Zhao et al. 2014).最近的研究表明含有碳水化合物结合域(carbohydrate binding module,CBM)的GEL4除了延伸β-1,3-葡聚糖外还可以建立分支,具有双重功能(Aimanianda et al. 2017).BGT1和BGT2基因也具有β-1,3-葡聚糖的分支功能,可以产生β-1,6-分支的β-1,3-葡聚糖,但它们的缺失没有引起细胞壁表型(Gastebois et al. 2010b),说明在细胞壁组装中它们并非重要基因.孢子萌发或菌丝分支生长中需要葡萄糖苷酶水解β-1,3-葡聚糖以增加细胞壁的可塑性.烟曲霉基因组含有多个β-1,3-葡聚糖酶,目前对其中eng1和eng2基因的功能分析发现它们的缺失未导致任何细胞壁缺陷(Mouyna et al. 2002;Hartl et al. 2011). ...
Deciphering the role of the chitin synthase families 1 and 2 in the in vivo and in vitro growth of Aspergillus fumigatus by multiple gene targeting deletion.
2
2014
... 几丁质是β-1,4-连接的N-乙酰葡萄糖胺线性聚合而成.已知几丁质由跨膜几丁质合成酶(Chitin synthase,CHS)以UDP- N-乙酰葡萄糖胺为前体合成.但几丁质合成机理尚不清楚,原因之一是丝状真菌中 CHS基因数量较多(Muszkieta et al. 2014).基于生物信息学分析烟曲霉有8个chs基因分属于两个家族.家族一的chsA、chsB和chsC对几丁质的合成贡献不大,但chsG单突变株及与chsA/B/C的多重突变株有分生孢子减少,菌丝形态改变、生长下降和几丁质合成酶活性降低等表型(Mellado et al. 1996;Mellado et al. 2003).家族二包括csmA、csmB、chsF和chsD 4个基因,4重敲除株表现出菌丝形态和孢子的变化,但几丁质含量几乎不受影响.敲除两个家族的4个基因(ΔcsmA/ΔcsmB/ ΔchsF/ΔchsG)后菌丝生长非常缓慢(Muszkieta et al. 2014),类似于fks1突变株,表明两个家族的几丁质合成酶共同协作参与烟曲霉几丁质的合成,也表明几丁质的合成对菌丝生长的重要性.构巢曲霉的细胞壁结构和烟曲霉的非常相似,但其几丁质合成酶的分类和功能与烟曲霉不同(Fujiwara et al. 2000;Tsuizaki et al. 2009),说明不同真菌中几丁质合成酶的功能根据细胞壁整体合成调控的需求有所不同. ...
... )后菌丝生长非常缓慢(Muszkieta et al. 2014),类似于fks1突变株,表明两个家族的几丁质合成酶共同协作参与烟曲霉几丁质的合成,也表明几丁质的合成对菌丝生长的重要性.构巢曲霉的细胞壁结构和烟曲霉的非常相似,但其几丁质合成酶的分类和功能与烟曲霉不同(Fujiwara et al. 2000;Tsuizaki et al. 2009),说明不同真菌中几丁质合成酶的功能根据细胞壁整体合成调控的需求有所不同. ...
Identification of Aspergillus fumigatus surface components that mediate interaction of conidia and hyphae with human platelets.
1
2015
... GAG是菌丝特有的组分,参与生物膜的形成(Fontaine et al. 2011),能促进菌丝与巨噬细胞、嗜中性粒细胞和血小板的结合(Rambach et al. 2015).另外还具有以下的直接免疫抑制作用:(1)通过NK细胞依赖性机制引发多形核中性粒细胞(polymorphonuclear neutrophil,PMN)凋亡并抑制PMN趋化性(Robinet et al. 2014);(2)介导对依赖于NADPH氧化酶的嗜中性粒细胞细胞外捕获的抗性(Lee et al. 2015).GM在菌丝生长过程中被分泌出来,是用于IA早期诊断的有效抗原之一(Foy et al. 2007).Bio-Rad开发的基于GM单克隆抗体的PlateliaTM检测试剂盒因其较好的特异性和灵敏性被广泛用于临床检测中.最近有报道称,Dectin-2与GM结合(Reedy et al. 2016),但这种C型凝集素的免疫功能尚不完全清楚. ...
Dectin-2 is a receptor for galactomannan. 7th Advances Against Aspergillosis Conference
1
2016
... GAG是菌丝特有的组分,参与生物膜的形成(Fontaine et al. 2011),能促进菌丝与巨噬细胞、嗜中性粒细胞和血小板的结合(Rambach et al. 2015).另外还具有以下的直接免疫抑制作用:(1)通过NK细胞依赖性机制引发多形核中性粒细胞(polymorphonuclear neutrophil,PMN)凋亡并抑制PMN趋化性(Robinet et al. 2014);(2)介导对依赖于NADPH氧化酶的嗜中性粒细胞细胞外捕获的抗性(Lee et al. 2015).GM在菌丝生长过程中被分泌出来,是用于IA早期诊断的有效抗原之一(Foy et al. 2007).Bio-Rad开发的基于GM单克隆抗体的PlateliaTM检测试剂盒因其较好的特异性和灵敏性被广泛用于临床检测中.最近有报道称,Dectin-2与GM结合(Reedy et al. 2016),但这种C型凝集素的免疫功能尚不完全清楚. ...
A polysaccharide virulence factor of a human fungal pathogen induces neutrophil apoptosis via NK cells
1
2014
... GAG是菌丝特有的组分,参与生物膜的形成(Fontaine et al. 2011),能促进菌丝与巨噬细胞、嗜中性粒细胞和血小板的结合(Rambach et al. 2015).另外还具有以下的直接免疫抑制作用:(1)通过NK细胞依赖性机制引发多形核中性粒细胞(polymorphonuclear neutrophil,PMN)凋亡并抑制PMN趋化性(Robinet et al. 2014);(2)介导对依赖于NADPH氧化酶的嗜中性粒细胞细胞外捕获的抗性(Lee et al. 2015).GM在菌丝生长过程中被分泌出来,是用于IA早期诊断的有效抗原之一(Foy et al. 2007).Bio-Rad开发的基于GM单克隆抗体的PlateliaTM检测试剂盒因其较好的特异性和灵敏性被广泛用于临床检测中.最近有报道称,Dectin-2与GM结合(Reedy et al. 2016),但这种C型凝集素的免疫功能尚不完全清楚. ...
Differential expression and extent of fungal/plant and fungal/bacterial chitinases of Aspergillus fumigatus.
1
2005
... 几丁质作为提供坚固性的细胞壁组分在形态发生中一直处于动态变化.几丁质酶水解β-1,4-糖苷键从而分解几丁质.烟曲霉有5个植物型、12个细菌型几丁质酶(Taib et al. 2005).植物型几丁质酶的五重敲除株与细菌型中AfChiB1的缺失都没有明显的表型(Jaques et al. 2003;Alcazar-Fuoli et al. 2011),说明不同类型的几丁质酶功能可以互补.另外几丁质脱乙酰酶和α-1,3-葡聚糖水解酶等在烟曲霉中也有多个冗余基因,利用遗传学技术分析这些酶的功能尚在进行中. ...
Myosin motor-like domain of the class VI chitin synthase CsmB is essential to its functions in Aspergillus nidulans.
1
2009
... 几丁质是β-1,4-连接的N-乙酰葡萄糖胺线性聚合而成.已知几丁质由跨膜几丁质合成酶(Chitin synthase,CHS)以UDP- N-乙酰葡萄糖胺为前体合成.但几丁质合成机理尚不清楚,原因之一是丝状真菌中 CHS基因数量较多(Muszkieta et al. 2014).基于生物信息学分析烟曲霉有8个chs基因分属于两个家族.家族一的chsA、chsB和chsC对几丁质的合成贡献不大,但chsG单突变株及与chsA/B/C的多重突变株有分生孢子减少,菌丝形态改变、生长下降和几丁质合成酶活性降低等表型(Mellado et al. 1996;Mellado et al. 2003).家族二包括csmA、csmB、chsF和chsD 4个基因,4重敲除株表现出菌丝形态和孢子的变化,但几丁质含量几乎不受影响.敲除两个家族的4个基因(ΔcsmA/ΔcsmB/ ΔchsF/ΔchsG)后菌丝生长非常缓慢(Muszkieta et al. 2014),类似于fks1突变株,表明两个家族的几丁质合成酶共同协作参与烟曲霉几丁质的合成,也表明几丁质的合成对菌丝生长的重要性.构巢曲霉的细胞壁结构和烟曲霉的非常相似,但其几丁质合成酶的分类和功能与烟曲霉不同(Fujiwara et al. 2000;Tsuizaki et al. 2009),说明不同真菌中几丁质合成酶的功能根据细胞壁整体合成调控的需求有所不同. ...
Immune interactions with pathogenic and commensal fungi: a two-way street
1
2015
... 细胞壁对真菌的生长和抵抗环境压力至关重要,同时它也是病原真菌与宿主免疫细胞最先接触的细胞器.先天性免疫反应对于有效及时地识别烟曲霉并诱导炎症网络清除非常重要(Becker et al. 2015).在免疫活性宿主中,肺泡巨噬细胞(AM)和嗜中性粒细胞每天需要清除数百至上千的烟曲霉分生孢子以防止肺部的炎性损伤(Underhill et al. 2015).肺泡巨噬细胞(alveolar macrophages,AM)是唯一可以进入肺泡腔的吞噬细胞而且是第一个接触吸入孢子的细胞.嗜中性粒细胞是烟曲霉的必需免疫效应细胞,因为严重嗜中性白血球减少症是侵袭性曲霉病发展的主要危险因素(Mircescu et al. 2009).微小的烟曲霉孢子被吸入宿主肺部时,细胞壁组分有的发挥激活宿主免疫反应的作用,有的则保护孢子并促进其在宿主中的定殖和生存(表1). ...
Fungal chitin dampens inflammation through IL-10 induction mediated by NOD2 and TLR9 activation
2
2014
... 几丁质长期以来在过敏和直接抗真菌响应方面发挥着重要作用.对几丁质的免疫应答是细胞类型特异性的,并随着几丁质颗粒的浓度和大小的不同而不同,在人和小鼠中有不一致的报道(Wagener et al. 2014).几丁质能诱导人几丁三糖糖苷酶产生,降解不溶性几丁质为可溶性壳寡糖,从而作为更强的免疫刺激剂(Wiesner et al. 2015)激活宿主免疫反应.在小鼠中,几丁质选择性诱导IL-10产生但不刺激促炎细胞因子(Wagener et al. 2014)的产生.不同宿主中免疫应答结果不一致的一个原因是目前没有几丁质与受体直接结合的证据.过去发现的几种几丁质受体(Levitz et al. 2015)诱导产生细胞因子的作用仍然存在争议. ...
... )激活宿主免疫反应.在小鼠中,几丁质选择性诱导IL-10产生但不刺激促炎细胞因子(Wagener et al. 2014)的产生.不同宿主中免疫应答结果不一致的一个原因是目前没有几丁质与受体直接结合的证据.过去发现的几种几丁质受体(Levitz et al. 2015)诱导产生细胞因子的作用仍然存在争议. ...
Chitin recognition via chitotriosidase promotes pathologic type-2 helper T cell responses to cryptococcal infection
1
2015
... 几丁质长期以来在过敏和直接抗真菌响应方面发挥着重要作用.对几丁质的免疫应答是细胞类型特异性的,并随着几丁质颗粒的浓度和大小的不同而不同,在人和小鼠中有不一致的报道(Wagener et al. 2014).几丁质能诱导人几丁三糖糖苷酶产生,降解不溶性几丁质为可溶性壳寡糖,从而作为更强的免疫刺激剂(Wiesner et al. 2015)激活宿主免疫反应.在小鼠中,几丁质选择性诱导IL-10产生但不刺激促炎细胞因子(Wagener et al. 2014)的产生.不同宿主中免疫应答结果不一致的一个原因是目前没有几丁质与受体直接结合的证据.过去发现的几种几丁质受体(Levitz et al. 2015)诱导产生细胞因子的作用仍然存在争议. ...
The Aspergillus fumigatus beta-1,3-glucanosyltransferase Gel7 plays a compensatory role in maintaining cell wall integrity under stress conditions.
1
2014
... 新合成的多糖链在细胞壁空间需要特定糖基转移酶和水解酶的共同作用进行重构、交联以产生细胞壁的三维网络结构,其中以β-1,3-葡聚糖的修饰最为多样.由葡聚糖合成酶复合体合成的线性β-1,3-葡聚糖需要经过延伸、分支和水解等过程才构成细胞骨架.葡聚糖的延伸是由糖基转移酶GEL家族负责,将水解后糖链的还原端转运到另一个葡聚糖分子的非还原端,从而延伸葡聚糖链.GEL属于CAZy数据库中的GH72家族,在烟曲霉中有7个成员,其中GEL4的缺失是致死的(Mouyna et al. 2005;Gastebois et al. 2010a),其他成员对葡聚糖的合成、形态发生、细胞壁完整性途径和致病力有不同程度的影响(Mouyna et al. 2005;Zhao et al. 2014).最近的研究表明含有碳水化合物结合域(carbohydrate binding module,CBM)的GEL4除了延伸β-1,3-葡聚糖外还可以建立分支,具有双重功能(Aimanianda et al. 2017).BGT1和BGT2基因也具有β-1,3-葡聚糖的分支功能,可以产生β-1,6-分支的β-1,3-葡聚糖,但它们的缺失没有引起细胞壁表型(Gastebois et al. 2010b),说明在细胞壁组装中它们并非重要基因.孢子萌发或菌丝分支生长中需要葡萄糖苷酶水解β-1,3-葡聚糖以增加细胞壁的可塑性.烟曲霉基因组含有多个β-1,3-葡聚糖酶,目前对其中eng1和eng2基因的功能分析发现它们的缺失未导致任何细胞壁缺陷(Mouyna et al. 2002;Hartl et al. 2011). ...

 , 金城
, 金城



{kind=link}
{kind=link}