
任春光
REN Chun-Guang
通讯作者:
收稿日期: 2017-07-17
接受日期: 2017-09-26
网络出版日期: 2018-02-22
版权声明: 2018 中国科学院微生物研究所《菌物学报》编辑部 版权所有
基金资助:
展开
摘要
为了鉴定冠突曲霉 veA基因缺失型与野生型的差异代谢物,寻找与 veA 基因产孢相关的代谢物,采用气相色谱-质谱联用(GC-MS)的代谢组学技术,分别收集2个菌株培养48h的菌丝体,经液氮研磨、超声萃取、三甲基氯硅烷衍生化后上机检测,将GC-MS检测的数据进行前处理和代谢物注释,并用统计学相关软件进行差异物筛选和相关性分析。结果显示,共鉴定99种代谢物,其中差异代谢物41种,野生型菌株中表达上调的显著差异代谢物有20种, veA缺失型菌株中表达上调的显著差异代谢物有21种,并预测差异代谢物的相关性,发现与623种代谢物产生相关性,其中313种呈正相关,310种呈负相关。筛选出来的差异代谢物涉及有机酸、氨基酸、碳水化合物、醇类、脂肪酸类等多种代谢物质,其中有机酸和氨基酸类代谢物占主导地位。本研究结果为进一步研究冠突曲霉代谢物与产孢的关联提供了一定的理论基础。
关键词:
Abstract
The metabolites of mycelial samples of ΔveA mutant and wild-type strain E4 of Aspergillus cristatus with 48 hour incubation period were identified by gas chromatography-mass spectrometry (GC-MS). A total of 99 metabolites were identified, including 41 significantly differential metabolites. Among them, 20 and 21 metabolites were much increased in the wild-type E4 and ΔveA, respectively. Statistical analysis revealed there were 623 individual metabolite having correlation, of which 313 were positive and 310 negative. These metabolites include organic acids, amino acids, carbohydrates, alcohols and oxylipin, while organic acids and amino acids predominate. This result lays the foundation for further study on the correlation between metabolite and sporulation in A. cristatus.
Keywords:
冠突曲霉 Aspergillus cristatus是茯砖茶在“发花”过程中形成的一种优势菌,俗称“金花”。茯砖茶是一种发酵茶,通常把“金花”的多少作为判断茯砖茶品质好坏的重要标志(刘作易等 1991)。由于“金花”具有抗氧化、促消化、降脂减肥、抑菌、抗癌等保健功效,使人们日益重视冠突曲霉的功能研究。
在真菌中, veA基因是一个全局性调节因子,该基因在许多丝状真菌中都发现既能够正向调控有性产孢也能够反向调控无性产孢,同时也能调节代谢物的变化,在有些真菌中是代谢物合成必须基因,敲除该基因对菌体形态和代谢物均有较大的影响(Yager et al. 1992;Calvo et al. 2008;马权等 2012;Hyoun et al. 2015)。形态建成与代谢物的合成是一个非常复杂的过程,它们之间互作引起形态、生理、分子水平上变化,涉及许多复杂因子的共同调控。目前国内对该菌的研究多集中于形态鉴定、药理活性物研究等方面。有关冠突曲霉代谢物与形态建成的关系还很少。该菌作为一种重要的工业微生物,探索冠突曲霉产孢过程中哪些代谢物发生了变化,以及这些代谢物变化的规律,研究其代谢物对产孢的影响,有助于控制合适的产孢方式和产孢数量,进而有效提高茯砖茶的质量和经济价值。
本实验室构建了 veA基因缺失型菌株,对 ∆veA菌株进行了显微观察,发现该突变体在麦芽酵母琼脂培养基上培养48h时, ∆veA缺失型只产生无性的分生孢子,而冠突曲霉野生型菌株在相同培养条件下只产生有性的子囊孢子。为了研究冠突曲霉形态变化与代谢物的关系,本实验拟采用代谢组学技术(GC-MS),收集野生型只产生子囊孢子时期和 ∆veA缺失型只产生无性的分生孢子时期的菌丝体,对其进行差异代谢组学研究。用一维方差分析(ANOVA)两组(E4和 veA)的显著性程度( P-value<0.05的为显著变化的代谢物),并结合 t检验( P<0.05)和倍性变化(>=1.5)来寻找差异的代谢物。寻找与 veA调控形态相关的差异代谢物,为进一步了解丝状真菌代谢物对产孢的调控机理打下基础。
1.1.1 供试菌株与培养基:冠突曲霉由贵州省农业生物技术重点实验室通过单孢分离获得并保存,菌株号为GZAAS 20.1005;遗传稳定的 ∆veA缺失型菌株由本实验室构建。5% MYA培养基(麦芽浸膏20g,蔗糖30g,酵母浸膏粉5g,琼脂15g,NaCl 5g)用于冠突曲霉野生型菌株(E4)和 ∆veA缺失菌株( veA)培养。
1.1.2 主要试剂及仪器:十九酸(0.2mg/mL)、1%三甲基氯硅烷、丙氨酸(10mmol/L)、色标正构烷烃标样C8-C20、C21-C40(Fluka Chemika,瑞士),甲醇(-20℃)等;主要仪器:Agilent 7890A/5975C气质联用仪(安捷伦,美国)、冷冻离心机、超净工作台、真空干燥机等。
样品的提取方法和衍生化是参照Ding(2009,2010)的研究。(1)将培养48h的冠突曲霉野生型菌株和 ∆veA缺失菌株用无菌刀刮取菌丝到5mL无菌离心管中,每个样品收集6个重复,共12个样品立即放在液氮中冷冻10min。(2)准确称取100mg用液氮研磨好的细胞置于2mL离心管中,在离心管中加入500μL甲醇(-20℃)和500μL ddH2O(4℃),漩涡30s,加入60μL十九酸(0.2mg/mL)作为内标和20μL同位素丙氨酸(10mmol/L),再次加入100mg玻璃珠漩涡1min,反复冻融3次;取出离心管,13 000× g 4℃离心10min,取上清液,离心浓缩干燥。加入60μL甲氧基溶液涡旋振荡30s,在37℃条件下反应2h。(3)最后加入60μL BSTFA试剂(含1%三甲基氯硅烷),37℃条件下反应90min。经过以上反应后移到进样瓶中室温静置2h,用于GC-MS分析。为了准确定性,引入烷烃保留指数,试剂为:色标正构烷烃标样C8-C20、C21-C40(Fluka Chemika,瑞士)。为了尽可能地避免系统误差,所有样品一批做完。
色谱条件:色谱柱HP-5MS毛细管柱(5% phenyl methyl silox:30m×250μm i.d.,0.25μm;agilent J&W scientific,Folsom,CA);分流进样,进样量1μL,分流比20:1。进样口温度280℃;离子源温度250℃;接口温度150℃。程序升温起始温度70℃;保持2min,以10℃/min升至300℃,保持5min。总运行时间为30min,载气为氦气,载气流速1mL/min。MS条件:电喷雾电离(ESI)源,全扫描方式,电子能量70eV;四极杆扫描范围m/z 35-780。
利用GC-MS预处理软件XCMS(www. bioconductor. org/)对Agilent 7890A/5975C气质联用仪检测获得的原始文件进行数据预处理,XCMS的最终结果导出至Excel进一步分析(Smith et al. 2006)。
对上述获得的保留时间、质核比及峰强的三维矩阵结合AMDIS程序进行代谢物的注释,注释所用数据库为国家标准与技术研究院(National Institute of Standards and Technology)和威利图书馆数据库(Wiley Registry),其中代谢物烷烃保留指数根据The Golm Metabolome Database(GMD)(http:// gmd.mpimp-golm.mpg.de/)提供的保留指数用于进一步的物质定性,同时大部分物质使用标准品进行进一步确认。
GC-MS数据经过XCMS软件处理的最终结果导出至Excel,保存为CSV格式,再将Excel数据导入SIMCA 11软件进行处理。采用默认的UV数据变换,首先用主成分分析(principal component analysis,PCA)进行无监督的数据分析,观察各族数据的聚类并去除离群样本;然后用偏最小二乘判别分析(partial least squares-discriminant analysis,PLS-DA)进行有监督的数据分析,并采用置换检验(permutation test)防止PLS-DA模型的过度拟合。利用一维方差分析(ANOVA)分别计算各组(E4和veA)的显著性程度,并进行假阳性矫正。最后取 P-value小于0.05为显著变化的差异代谢物。并结合 t检验( P<0.05)和倍性变化(>=1.5)来寻找差异的代谢物, t统计学检验用R 3.3.0(www.r-project.org)进行。采用皮尔森相关系数(Pearson’s product-moment correlation),计算每个相关性的 P值, P值用false discovery raete(FDR)进行矫正,关联性矩阵用R语言的cor函数和cor.test进行计算。并根据(|r|>0.8和FDR<0.05)筛选显著关联性,根据相关系数绘制代谢物关联热图。
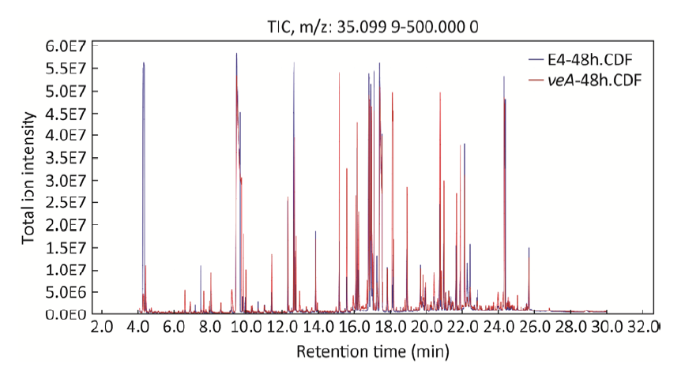
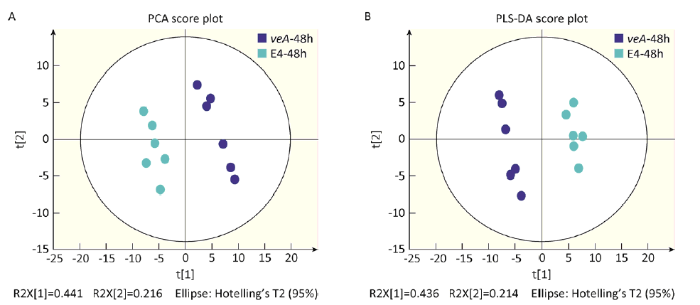
经过GC-MS检测以后,得到冠突曲霉野生型和 ∆veA缺失型的总离子流图(图1)。冠突曲霉野生型和 ∆veA缺失菌株两个样品的峰及保留时间几乎是相同的,其整体的差异主要体现在峰高和峰面积上差异较显著。对获得的保留时间、质核比及峰强的矩阵进行代谢物的注释,最终获得99个可以被数据库注释的代谢物(表1),大量的物质由于信号低或者数据库没有收录而没有被注释。为了确定2个样本之间是否具有差异,采用了主成分分析(PCA)、偏最小二乘法分析(PLS-DA)来区分(图2),在图2中的A和B图上的每个点表示一个对应的样本,从PCA得分图可以看出GC-MS分析所得原始数据在PCI和PC2 2种主成分中都能清楚区分开,PLS-DA也能够清楚区分开,通过2个模型相互论证说明冠突曲霉野生型与 ∆veA缺失型2样本差异明显。通过图的形式得到的分析结果将更加直观和可靠,且具有统计学意义。
图1 冠突曲霉 ∆veA缺失型和野生型样本的GC-MS总离子流图
E4:蓝色峰表示野生型; veA:红色峰表示 ∆veA缺失型
Fig. 1 Total ion current (TIC) chromatogram of ∆veA mutant and wild type strain
E4 of Aspergillus cristatus with 48h incubation period. E4: The wild type (blue peak); veA: ∆veA mutant group (red peak).
表1 GC-MS检测冠突曲霉 ∆veA缺失型和野生型孢内代谢物
Table 1 The intracellular metabolites identified by GC-MS of ∆veA mutant and wild type E4 of Aspergillus cristatus in 48h of incubation
| 物质名称 Name | 保留指数 RI | 质荷比 m/z | 保留时间 RT | ∆veA-48h vs E4-48h | |
|---|---|---|---|---|---|
| VIP | P-value | ||||
| Pyruvic acid | 1057.3 | 88.996 | 365.556 | 0.992 | 0.026 |
| Lactic acid+ | 1068.7 | 190.029 | 375.828 | 0.637 | 0.186 |
| Glycolic acid | 1082.6 | 177.003 | 388.957 | 0.675 | 0.159 |
| Alanine+ | 1109.7 | 116.000 | 414.018 | 0.159 | 0.752 |
| 3-hydroxypropanoic acid | 1150.8 | 176.994 | 451.248 | 0.052 | 0.918 |
| Monomethylphosphate | 1186.9 | 241.000 | 484.196 | 0.254 | 0.612 |
| Malonic acid+ | 1211.2 | 233.000 | 505.48 | 0.911 | 0.045 |
| 2-oxoisocaproic acid | 1222 | 200.066 | 514.791 | 1.044 | 0.017* |
| Valine+ | 1224.3 | 144.100 | 517.0325 | 0.380 | 0.444 |
| 4-hydroxy-butyric acid+ | 1241.1 | 116.973 | 531.563 | 1.071 | 0.013* |
| Benzoic acid+ | 1251.7 | 178.993 | 540.413 | 0.695 | 0.145 |
| Urea+ | 1260.5 | 189.009 | 552.317 | 0.890 | 0.052 |
| Ethanolamine | 1275.2 | 174.095 | 560.518 | 0.637 | 0.186 |
| Leucine+ | 1281.6 | 158.100 | 565.855 | 0.442 | 0.371 |
| Glycerol+ | 1295.8 | 205.000 | 578.999 | 1.228 | 0.002* |
| Phosphoric acid | 1297.7 | 299.098 | 580.558 | 0.357 | 0.473 |
| Threonine+ | 1307.5 | 117.000 | 587.979 | 1.292 | 0.001* |
| Proline+ | 1310.4 | 142.075 | 590.1685 | 0.162 | 0.747 |
| Glycine+ | 1319 | 174.099 | 597.6325 | 0.137 | 0.786 |
| Succinic acid+ | 1320.4 | 247.099 | 598.405 | 1.198 | 0.003* |
| Glyceric acid | 1343.1 | 292.100 | 616.84 | 0.818 | 0.079 |
| Uracil | 1348.1 | 99.000 | 621.051 | 1.249 | 0.002* |
| Fumaric acid+ | 1351.2 | 245.000 | 623.5765 | 1.399 | 0.000* |
| Itaconic acid+ | 1359.1 | 259.024 | 630.0095 | 1.297 | 0.001* |
| Homoserine | 1364.1 | 146.000 | 634.253 | 0.265 | 0.597 |
| Serine+ | 1372.1 | 204.100 | 640.5865 | 0.257 | 0.608 |
| Mevalonic acid-1,5-lactone | 1376.4 | 145.000 | 644.4185 | 1.390 | 0.000* |
| 2-methyl-1,3-butanediol | 1390.9 | 117.000 | 655.672 | 1.138 | 0.007* |
| Allothreonine | 1399.2 | 117.000 | 662.574 | 0.214 | 0.670 |
| Glutaric acid+ | 1408 | 186.000 | 610.2085 | 1.479 | 0.000* |
| N,N-di-(2-hydroxyethyl)-methanamine | 1430.6 | 160.000 | 686.158 | 1.143 | 0.006* |
| Beta-alanine+ | 1436.1 | 174.010 | 686.804 | 0.239 | 0.633 |
| Erythronic acid-1,4-lactone | 1443.3 | 233.065 | 696.1045 | 1.111 | 0.009* |
| Nicotinamide+ | 1485.9 | 179.000 | 729.7865 | 1.503 | 0.000* |
| Malic acid+ | 1501.7 | 233.100 | 740.0755 | 0.721 | 0.129 |
| Erythritol | 1519.2 | 217.100 | 752.906 | 1.358 | 0.000* |
| Threitol | 1530 | 217.100 | 759.329 | 1.407 | 0.000* |
| Pyroglutamic acid+ | 1533.6 | 156.000 | 763.822 | 0.626 | 0.195 |
| Aspartic acid+ | 1534.4 | 100.000 | 764.139 | 0.676 | 0.158 |
| N-acetylglutamic acid | 1539.8 | 84.000 | 767.3565 | 0.748 | 0.113 |
| 2,4,6-tri-tert-butylbenzenethiol | 1552.7 | 263.186 | 777.298 | 1.121 | 0.008* |
| Phenylalanine+ | 1554.6 | 120.000 | 778.5835 | 1.251 | 0.002* |
| Threonic acid+ | 1563.7 | 292.100 | 785.317 | 1.281 | 0.001* |
| Citramalic acid | 1569.9 | 247.100 | 789.797 | 1.343 | 0.000* |
| 6-hydroxy-3-pyridinecarboxylic acid | 1577.8 | 268.071 | 795.435 | 1.112 | 0.009* |
| Tetronic acid | 1580.1 | 292.100 | 797.1945 | 0.883 | 0.054 |
| 2-hydroxyglutaric acid+ | 1586 | 129.000 | 801.084 | 1.346 | 0.000* |
| 3-hydroxy-3-methylglutaric acid | 1614.2 | 247.088 | 820.943 | 0.912 | 0.045 |
| Glutamic acid+ | 1632.2 | 246.100 | 832.864 | 1.000 | 0.033 * |
| 4-hydroxybenzoic acid | 1633.3 | 267.084 | 834.121 | 0.858 | 0.063 |
| Glucopyranose | 1668.9 | 204.093 | 862.339 | 0.660 | 0.169 |
| 2-ketogluconic acid | 1675 | 204.093 | 862.339 | 0.660 | 0.169 |
| Arabinose+ | 1687.7 | 103.000 | 871.006 | 0.472 | 0.338 |
| Ribose+ | 1703 | 103.000 | 881.274 | 0.862 | 0.062 |
| Lysine+ | 1718 | 84.016 | 890.908 | 0.701 | 0.142 |
| Arabinitol+ | 1747.4 | 217.100 | 910.159 | 1.545 | 0.000* |
| Glycerol-2-phosphate | 1749 | 299.085 | 911.122 | 1.379 | 0.000* |
| Aconitic acid | 1759 | 229.091 | 917.662 | 1.034 | 0.018* |
| 2-desoxy-pentos-3-ylose dimethoxyamine | 1777.3 | 231.100 | 929.408 | 0.823 | 0.077 |
| Glycerol-3-phosphate | 1784.7 | 299.100 | 934.438 | 1.478 | 0.000* |
| alpha-d-methylfructofuranoside | 1801.5 | 217.100 | 944.501 | 0.132 | 0.793 |
| Ribonic acid | 1809.5 | 292.100 | 950.272 | 1.411 | 0.000* |
| 3-deoxyglucitol | 1813.2 | 103.000 | 952.845 | 0.283 | 0.571 |
| Hypoxanthine | 1818.6 | 265.100 | 956.121 | 1.185 | 0.004* |
| Citric acid+ | 1844.1 | 273.100 | 970.536 | 0.176 | 0.726 |
| 2-methyl-citric acid | 1862 | 287.095 | 982.415 | 1.468 | 0.000* |
| O-methyl-inositol | 1864 | 217.100 | 983.986 | 1.430 | 0.000* |
| Diisobutyl-phthalate | 1874.4 | 149.000 | 990.058 | 0.738 | 0.119 |
| Tyrosine+ | 1897.1 | 179.100 | 1004.200 | 1.309 | 0.001* |
| Fructose+ | 1906.5 | 103.059 | 1012.806 | 1.191 | 0.004* |
| Gluconic acid-1,5-lactone | 1915.6 | 220.100 | 1014.068 | 1.523 | 0.000* |
| Glucose+ | 1934.2 | 319.193 | 1026.326 | 1.495 | 0.000* |
| Galactose+ | 1952.6 | 319.191 | 1036.921 | 1.071 | 0.013* |
| Mannitol+ | 1967 | 217.100 | 1045.580 | 0.608 | 0.209 |
| Galactonic acid | 1994 | 292.000 | 1060.689 | 1.103 | 0.010* |
| Pantothenic acid | 2014.2 | 103.000 | 1070.657 | 0.906 | 0.047 |
| Galactaric acid | 2041.1 | 333.100 | 1087.941 | 1.552 | 0.000* |
| Hexadecanoic acid | 2047.9 | 117.000 | 1091.156 | 0.907 | 0.047 |
| Glucaric acid | 2058.2 | 292.100 | 1095.979 | 1.041 | 0.017* |
| 9,12-octadecadienoic acid (z,z)-, methyl ester | 2094.8 | 81.005 | 1118.112 | 0.184 | 0.715 |
| 9-octadecenoic acid (z)-, methyl ester | 2100.3 | 83.000 | 1121.075 | 0.403 | 0.416 |
| N-acetyl glucosamine | 2116.8 | 205.100 | 1129.701 | 1.114 | 0.009* |
| Myo-inositol+ | 2126.8 | 305.100 | 1135.123 | 0.917 | 0.044 |
| Uric acid+ | 2128.4 | 441.200 | 1136.404 | 1.080 | 0.012* |
| Guanine+ | 2144.8 | 352.136 | 1145.067 | 0.564 | 0.247 |
| 9,12-(z,z)-octadecadienoic acid+ | 2213.3 | 81.003 | 1181.329 | 1.205 | 0.003* |
| 9-(z)-octadecenoic acid+ | 2218.1 | 117.000 | 1183.579 | 1.374 | 0.000* |
| Octadecanoic acid+ | 2243.4 | 117.000 | 1196.418 | 1.000 | 0.033* |
| Fructose-6-phosphate+ | 2356.5 | 299.088 | 1249.244 | 0.026 | 0.959 |
| Sorbitol-6-phosphate | 2403.1 | 299.095 | 1277.587 | 1.104 | 0.010* |
| Insitol-1-phosphate | 2468.7 | 318.117 | 1307.646 | 0.192 | 0.702 |
| Uridine+ | 2477.4 | 217.100 | 1313.227 | 1.344 | 0.000* |
| Inosine+ | 2603 | 217.100 | 1370.676 | 1.474 | 0.000* |
| Adenosine+ | 2621.3 | 245.087 | 1378.718 | 0.652 | 0.175 |
| Guanosine+ | 2799 | 245.100 | 1454.609 | 0.223 | 0.657 |
| Trehalose+ | 2809.8 | 204.100 | 1460.215 | 1.494 | 0.000* |
| Gentibiose | 2889.5 | 361.200 | 1489.088 | 0.631 | 0.191 |
| Melibiose | 2944 | 204.100 | 1515.389 | 0.843 | 0.069 |
| Galactinol | 2996.6 | 204.100 | 1539.958 | 1.041 | 0.017* |
图2 冠突曲霉 ∆veA缺失型与野生型PCA(A)和PLS-DA(B)得分图
Fig. 2 Principal component analysis (PCA) plot (A) and PLS-DA analysis plot (B) of ΔveA mutant and wild-type E4 of Aspergillus cristatus in 48h of incubation. E4: Wild-type; veA: ∆veA mutant.
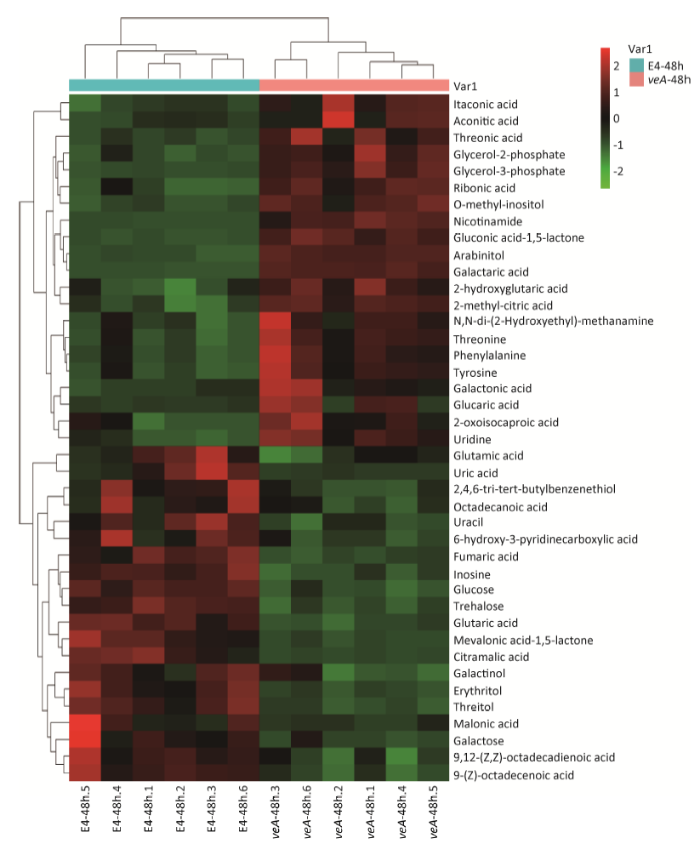
冠突曲霉 ∆veA缺失型与野生型进行比较。用偏最小二乘法判别分析,VIP>1和 P<0.05的视为显著差异。结果表明,冠突曲霉 ∆veA缺失型与野生型之间有48种代谢物差异显著(表1)。为了进一步论证寻找差异代谢物的准确性,随后通过聚类热图用 P<0.05和倍性变化≥1.5进行筛选,发现有41种差异代谢物(图3)。虽然用2种不同的标准寻找差异代谢物,但差异代谢物最终的结论几乎是相同的,即有40种代谢物完全相同(差异代谢物相同率83%)。用2种筛选方法共同确定差异代谢物,最终确定冠突曲霉 ∆veA缺失型与野生型有41种差异代谢物。聚类热图分析表明,相对含量上调的有21种,下调的有20种(图3)。这些显著差异代谢物包括17种有机酸、7种氨基酸和氨基酸衍生物、5种碳水化合物、4种醇类物质、3种脂肪酸物质、2种磷酸盐、2种胺类物质和1种酚类物质,其中有机酸占主导地位。
图3 冠突曲霉 ∆veA缺失型与野生型显著差异代谢物聚类热图
红色代表代谢物相对量增加,绿色代表代谢物相对量降低
Fig. 3 Hierarchical cluster analysis (HCA) of intracellular metabolites of ΔveA mutant and wild-type E4 of Aspergillus cristatus.
Red squares in the heat map indicate increases of intracellular metabolite concentration, while green squares indicate decreases of intracellular metabolite concentration.
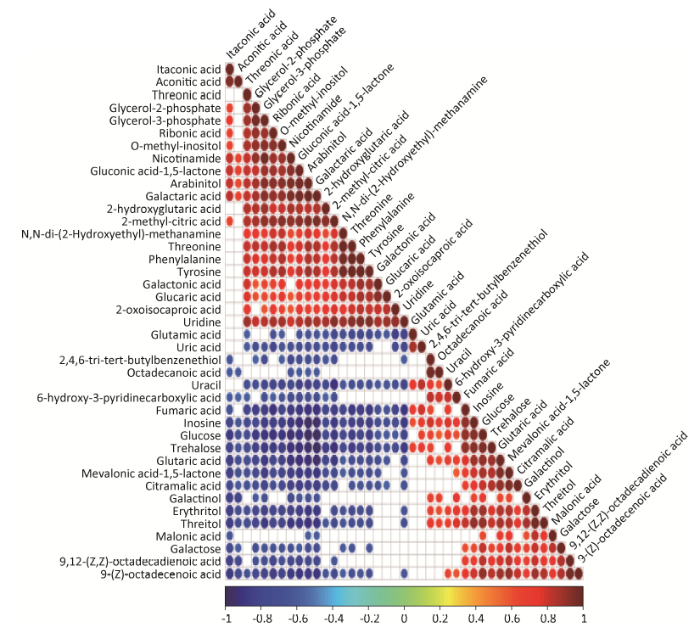
采用相关系数分析法,分析已经确定的41种差异代谢物的相关性,有623种代谢物相关,其中313种物质呈正相关,310种物质呈负相关(图4),在正相关性中,有机酸占主导地位,其次是氨基酸、碳水化合物、醇类、磷酸盐和脂肪酸类,最后是胺类和酚类代谢物。在负相关性中,有机酸占主导地位,其次是碳水化合物、氨基酸、脂肪酸、醇类、最后是酚类代谢物。结果表明冠突曲霉形态变化过程中,有机酸、氨基酸、碳水化合物、醇类、磷酸盐和胺类代谢物可能对冠突曲霉产孢均起到重要作用,其中有机酸和氨基酸占主导地位。
图4 冠突曲霉 ∆veA缺失型与野生型差异代谢物相关性分析
红色表示完全的正相关;蓝色表示完全的负相关;图中空白部分为相关性统计检验 P>0.05;有颜色标记部分为 P<0.05,视为代谢物显著相关
Fig. 4 Metabolite-metabolite correlation analysis of ΔveA mutant and wild-type E4 of Aspergillus cristatus.
Positive correlation is shown in red and negative correlation in blue ( P<0.05). Blank squares: P>0.05. Marked with red or blue ( P<0.05) are the significant metabolite-metabolite correlations.
代谢组学(metabolomics/metabonomics)是继基因组学、转录组学、蛋白质组学提出的一门新学科。目前寻找差异代谢物主要用两种分析手段,GC-MS和HPLC-MS。GC-MS方法克服了HPLC-MS数据库不全,检测出来的代谢物无法鉴定等缺点。GC-MS数据库较全面,容易鉴定相关代谢物,是目前用于寻找差异代谢物研究最有效、检测效率高、检测灵敏、应用最广泛的方法之一,是研究复杂混合物分析的主要定性和定量手段之一。目前为止,利用该技术寻找差异代谢物的报道已有很多,Roze et al.(2010)运用此技术研究了寄生曲霉中野生型与 veA缺失型差异代谢物,发现了支链氨基酸和乙醇新的代谢途径,孢内挥发性代谢产生依赖 veA调控。Ding et al.(2010)采用GC-MS技术研究了不同酵母菌株在乙醇的胁迫下产生了一些差异代谢物。Han et al.(2012)采用GC-MS技术研究了白色念珠菌形态转变过程中代谢物的变化,鉴定了75种代谢物,发现19种差异代谢物。
本实验采用GC-MS技术,以野生型菌株产生的子囊孢子和 veA缺失菌株产生的分生孢子为材料,筛选出一些差异代谢物。这些差异代谢物涉及到有机酸类代谢、氨基酸类代谢、碳水化合物代谢、醇类代谢、磷酸盐类代谢等各个方面。本试验得到的差异代谢物衣康酸(itaconic acid)是属于有机酸类代谢物,它是一种重要的工业物质。有研究发现衣康酸含量对土曲霉菌丝生长及孢子形成有显著影响,观察菌丝生长率和孢子大小可筛选出高产衣康酸的突变菌株(Shin et al. 2011)。
在野生型中酪氨酸(tyrosine)和苯丙氨酸(phenylalanine)的相对含量显著下调。Han(2012)等科学家已报道其代谢物含量变化是白色念珠菌从菌丝转换成孢子的关键影响因子。此外,试验筛选到的葡萄糖,已有研究证明葡萄糖浓度增加,能够促进构巢曲霉闭囊壳数量增加,乳糖含量增加有利于构巢曲霉闭囊壳的发育(Han et al. 2003)。试验筛选到的差异代谢与已经报道形态相关的物质进行比较,仅有少部分被报道,其余大部分代谢物与产孢相关研究还未见报到,例如有机酸类物质:苏糖酸(threonic acid)、尿酸(uric acid)、粘酸(galactric acid)等,氨基酸类物质:苏氨酸(threonine)、谷氨基酸(glutamic acid)等,醇类物质:苏糖醇(threitol)、赤藓糖醇(erythritol)等,糖类物质:海藻糖(trehalose),脂肪酸类物质:9,12-(Z,Z)-octadecadienoic acid、9-(Z)-octadecenoic acid等物质。这些结果显示尚有很多参与真菌形态建成的物质需要更进一步研究。
在此之前,不断有研究报道,真菌形态建成受多种类代谢影响,例如醇类:金合欢醇、酪醇、苯乙基乙醇、甘露醇等物质,其中部分醇类物质(甘露醇)是曲霉形态建成的必须物质,甘油浓度增加有利于构巢曲霉闭囊壳的发育(Han et al. 2003;Chen et al. 2004;Chen & Fink 2006;Nickerson et al. 2006;Wyatt et al. 2014)。氨基酸类物质:酪氨酸、脯氨酸、苯丙氨酸等;脂肪酸类:豆蔻酸;盐类有苹果酸盐和琥珀酸盐等,这些代谢物均对白色念珠菌形态转换起关键作用(Han et al. 2012)。脂肪酸具有多种生物学功能,例如影响菌丝生长、生物膜的形成、调控形态转变、影响群体感应、影响有性孢子和无性孢子的比例(Kinderlerer 1993;赵人俊和郑幼霞 1995;Chen et al. 2004;Dimitrios & Nancy 2007;Christensen & Kolomiets 2011;Han et al. 2011)。Mazur(1991)已经证明了氧化脂类物质(psiBa)能够促进曲霉菌有性孢子发育。 veA基因既能正向调控有性发育,也能反向调控无性发育,在完全相同的培养条件下培养48h,野生型菌株产生有性孢子, ∆veA缺失型菌株产生无性孢子,因此,可以推测,在 ∆veA缺失型菌株中,某些物质增加或减少有利于无性孢子的产生。
综上所述,本试验筛选的有机酸、氨基酸、糖类代谢物、醇类物质和磷酸盐等受 veA基因调控,其中有机酸和氨基酸类代谢物对冠突曲霉产孢起重要作用。这些代谢组信息为我们继续研究关键代谢物参与的代谢途径、代谢途径上关键基因的功能及其在冠突曲霉产孢方面所起的作用提供了很好的借鉴。同时通过了解冠突曲霉产孢过程中代谢物的变化规律,为后续理性优化添加关键代谢物,为提高当下茯砖茶质量及有益物质积累提供新思路。
致谢:本研究在实验方案设计等诸多方面得到了中国科学院微生物研究所尹文兵研究员和贵州大学生命科学学院葛永怡博士的大力支持,在此表示衷心感谢!
| [1] |
The veA regulatory system and its role in morphological and chemical development in fungi .
|
| [2] |
Feedback control of morphogenesis in fungi by aromatic alcohols . |
| [3] |
Tyrosol is a quorum-sensing molecule in Candida albicans . |
| [4] |
The lipid language of plant-fungal interactions . |
| [5] |
Oxylipins as developmental and host-ungal communication signals . |
| [6] |
Comparative metabolomic analysis on industrial continuous and batch ethanol fermentation processes by GC-TOF-MS .
|
| [7] |
Metabolome analysis of differential responses of diploid and haploid yeast to ethanol stress .
|
| [8] |
Environmental factors affecting development of Aspergillus nidulans . |
| [9] |
The metabolic basis of Candida albicans morphogenesis and quorum sensing . |
| [10] |
Metabolome analysis during the morphological transition of Candida albicans .
|
| [11] |
The veA gene is necessary for the negative regulation of the veA expression in Aspergillus nidulans .
|
| [12] |
Fungal strategies for detoxification of medium chain fatty acids . |
| [13] |
Study of conditions of sporogenesis of Aspergillus chevalieri var. intermedius in “Fuzhuan” tea . |
| [14] |
Research progress of veA gene in filamentous fungi .
|
| [15] |
Structure and synthesis of sporogenic psi factors from Aspergillus nidulans .
|
| [16] |
Quorum sensing in dimorphic fungi: farnesol and beyond . |
| [17] |
Volatile profiling reveals intracellular metabolic changes in Aspergillus parasiticus: veA regulates branched chain amino acid and ethanol metabolism .
|
| [18] |
Development of an efficient screening strategy for rapid selection of high-yielding mutants of itaconic acid biosynthesized by fungal cells of Asperillus terreus .
|
| [19] |
XCMS: processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification . |
| [20] |
Mannitol is essential for the development of stress-resistant ascospores in Neosartorya fischeri (Aspergillus fischeri) . |
| [21] |
Early developmental events during asexual and sexual sporulation in Aspergillus nidulans .
|
| [22] |
Studies on the production of lipid in filamentous fungus Mortierella sp . |
| [23] |
茯砖茶“金花”菌—— 谢瓦氏曲霉间型变种的孢子产生条件 .
|
| [24] |
丝状真菌 veA基因的研究进展 .
|
| [25] |
丝状真菌被孢霉产生油脂的研究 .
|



/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}