Aspergillus cell wall melanin blocks LC3-associated phagocytosis to promote pathogenicity
1
2016
... 次级代谢产物有利于真菌在激烈的竞争环境中生存,已经有大量研究表明真菌次级代谢产物在真菌与其他微生物、真菌与昆虫以及真菌与植物之间都起到非常重要的作用(Rohlfs 2015;Zeilinger et al. 2016;Scherlach & Hertweck 2017).如上文提到的次级代谢产物黑色素,不仅可以保护真菌孢子免受紫外线的伤害,而且可以作为毒力因子.有研究表明通过各种机制影响巨噬细胞在抗真菌免疫过程中的识别和吞噬等环节,降低其抗真菌效应,在烟曲霉感染中,黑色素可通过覆盖细胞壁表面的β-甘露糖,影响巨噬细胞PRRs对烟曲霉识别,帮助烟曲霉在巨噬细胞内存活(Akoumianaki et al. 2016);通过CRISPR/Cas9敲除皮炎外瓶霉的黑色素合成关键酶PKS1基因,也发现白化株对人离体皮肤组织的侵袭力较色素株下降,这表明黑色素能够增强皮炎外瓶霉的侵袭力(Poyntner et al. 2018).在真菌与细菌的研究中,真菌与细菌通过次级代谢产物相互作用中的例子有很多,细菌青枯雷尔氏菌Ralstonia solanacearum分泌脂肽ralsolamycin,其诱导真菌中的厚垣孢子形成和镰刀菌属中的bikaverin基因簇的表达,进而可减少细菌进入和生长(Spraker et al. 2016);研究发现伊氏杀线真菌胞内存在内生细菌,其合成的次级代谢产物将有助于伊氏杀线真菌在宿主线虫内定殖,增加伊氏杀线菌在生态竞争中的适应性,以便更好地捕杀线虫,减少松树枯萎病(王瑞珍 2017).在防御昆虫的过程中,真菌往往产生一些有毒的次级代谢产物来保证自身的安全.例如:白僵菌Beauveria bassiana可以通过其次级代谢产物白僵菌素杀死害虫,而且还用聚酮类卵磷脂毒化尸体,以限制其他微生物对其食物的竞争(Fan et al. 2017);同样的,黄曲霉产生的有毒次级代谢产物黄曲霉毒素对昆虫也有一定的杀伤力,研究发现,在黄曲霉毒素丰富的环境中,在与昆虫竞争的试验中,真菌的适应性提高了26倍,而黄曲霉产毒菌株显示出更高的适应性(Keller 2019). ...
Distinct roles for VeA and LaeA in development and pathogenesis of Aspergillus flavus
1
2009
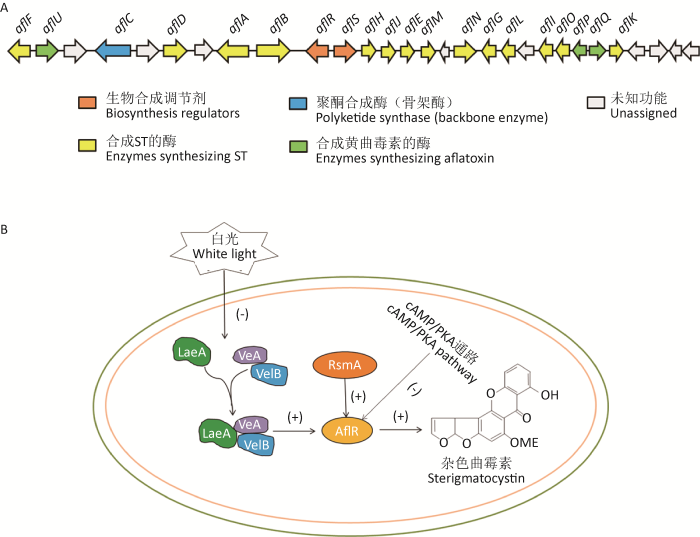
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Low-volume toolbox for the discovery of immunosuppressive fungal secondary metabolites
1
2013
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Regulation of fungal secondary metabolism
2
2013
... 真菌在环境中无处不在,在植物、动物和人类的健康以及广泛的生态系统功能中都发挥着重要作用(Xu 2016).它们能产生各种各样的次级代谢产物,而次级代谢产物对其自身的正常生长不是必需的化合物,但其在真菌生态学中作为适应因子扮演着重要的角色,例如紫外线防护,并且能够通过在生态相互作用中作为防御化合物或信号分子起作用(Brakhage 2013;Keller 2019).不仅如此,真菌产生的这些次级代谢产物在人类生活中具有重要的应用,自弗莱明从青霉属Penicillium中发现青霉素并广泛应用以来,真菌次级代谢产物成为药物分子的重要来源(Raja et al. 2017),例如,他汀类、环孢菌素和霉酚酸等已经开始大规模地用于延长人类生命(Keller 2019).此外,真菌产生的次级代谢产物也表现出对人有害的特性,例如,黄曲霉毒素(AF)、环匹阿尼酸和黄曲霉震颤素等.由于次级代谢产物的重要性,所以对次级代谢产物的合成调控研究已经成为热点. ...
... 次级代谢产物的合成是一个复杂过程,需要多个基因参与调控,基因组挖掘工作表明,真菌产生次级代谢产物的能力被大大低估,因为许多真菌次级代谢产物生物合成基因簇在标准培养条件下是沉默的(Brakhage 2013).类似于原核生物的操纵子,在真菌的基因组中也出现了彼此功能相关的非同源基因成簇存在的现象.这些基因形成基因簇,可参与多种次级代谢途径(Osbourn 2010).真菌次级代谢基因簇由复杂的调节网络控制,其全局调控主要通过全局调控因子实现,而通路特异性调节由它们调节簇内的基因编码的转录因子介导.本文主要根据近年来对次级代谢产物的研究,综合论述了真菌次级代谢产物的药用价值以及在竞争、防御和发育过程中的生态功能,此外,还描述了次级代谢物生物合成基因簇的转录和表观遗传调控,旨在更加深刻地理解次级代谢产物的合成调控. ...
Lae1 regulates expression of multiple secondary metabolite gene clusters in Fusarium verticillioides
1
2012
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Genetic connection between fatty acid metabolism and sporulation in Aspergillus nidulans
1
2001
... 真菌产生的次级代谢产物与真菌的发育有着密切联系,上文中已经提到真菌黑色素不仅能够抵抗紫外线辐射,而且能够结合和隔离非特异性肽和化合物,产生保护细胞的物理屏障,为真菌孢子提供保护,使其免受极端环境或昆虫捕食.除此以外,研究表明,土曲霉产生的丁内酯,能在真菌中提高菌丝的分枝、孢子形成(Schimmel et al. 1998);在构巢曲霉中,内源性油酸和亚麻油酸衍生物能调控形成无性和有性孢子的比例(Calvo et al. 2001);镰刀菌属的色素形成需要镰刀菌素(Lena et al. 2012);神经孢菌素neurosporin A可以阻止食虫动物以粗糙脉孢菌孢子为食(Zhao et al. 2017).事实上,细菌的次级代谢产物也会对真菌发育产生影响,次级代谢产物可以作为种间信号,通过氧化应激调节影响真菌发育.在共培养实验中,铜绿假单胞菌次级代谢产物吩嗪对烟曲霉发育有不同的调节作用,其中高浓度的吩嗪具有抗真菌作用,但中等浓度会诱导真菌产孢(He et al. 2015). ...
Anticancer activity of an endophytic fungus Trichoderma sp. from a horse
0
2017
Functions of fungal melanin beyond virulence
2
2017
... 自古以来,人们就将真菌作为药材使用,《神农本草经》和《本草纲目》等书中就有记载,例如茯苓、灵芝等真菌药物.不仅如此,其产生的次级代谢产物在抗菌、抗心血管疾病、抗肿瘤和抗病毒等方面表现出不同的活性,具有重要的药用价值(Wu et al. 2019),其中一些次级代谢产物已经被开发成药物,例如:具有抗菌活性的药物青霉素、灰黄霉素就是从自然界中常见的青霉属Penicillium中分离得到,而卡泊芬净是由Glarea lozoyensis的发酵产物合成的;免疫抑制剂环孢菌素来自Trichoderma和Tolypocladium属;治疗心血管疾病的药物洛伐他汀类药物来自Aspergillus等属.还有许多次级代谢产物还没有被研发成药物,但是具有重要的药物活性,如陈旭等(2017)从追风伞植物中分离得一株木霉属内生真菌可以产甾体化合物,发现其能抑制两种癌细胞的增殖;孔阳(2019)从白花夹竹桃Nerium indicum mill. cv Paihua中分离出3株具有强抗菌活性的内生真菌,实验结果显示其次级代谢产物不仅对人前列腺癌细胞PC3、人肺癌细胞A549和人乳腺癌细胞MCF-7有良好的抑制效果,而且有一定的抗糖尿病活性;大型真菌Hericium alpestre产生的次级代谢产物——4-羟基-2-吡啶酮类生物碱sambutoxin具有良好的抗肿瘤作用(李璐宁 2019);周岳(2018)从海洋真菌Aspergillus versicolor LZD4403中分离得到的次级代谢产物曲林菌素可以通过抑制巨噬细胞炎症,进而发挥抑制巨噬细胞泡沫化作用,从而发挥抗动脉粥样硬化的作用;近年来研究发现真菌次级代谢产物黑色素在体外对艾滋病病毒有显著的抑制作用,这使天然黑色素有望成为一种新的抗艾滋病药物(Cordero & Casadevall 2017).以上研究均表明,真菌次级代谢产物在人类医疗健康方面具有重大价值,因此,了解真菌次级代谢产物的功能和合成调控具有重大意义. ...
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Regulatory cascade and biological activity of Beauveria bassiana oospore in that limits bacterial growth after host death
1
2017
... 次级代谢产物有利于真菌在激烈的竞争环境中生存,已经有大量研究表明真菌次级代谢产物在真菌与其他微生物、真菌与昆虫以及真菌与植物之间都起到非常重要的作用(Rohlfs 2015;Zeilinger et al. 2016;Scherlach & Hertweck 2017).如上文提到的次级代谢产物黑色素,不仅可以保护真菌孢子免受紫外线的伤害,而且可以作为毒力因子.有研究表明通过各种机制影响巨噬细胞在抗真菌免疫过程中的识别和吞噬等环节,降低其抗真菌效应,在烟曲霉感染中,黑色素可通过覆盖细胞壁表面的β-甘露糖,影响巨噬细胞PRRs对烟曲霉识别,帮助烟曲霉在巨噬细胞内存活(Akoumianaki et al. 2016);通过CRISPR/Cas9敲除皮炎外瓶霉的黑色素合成关键酶PKS1基因,也发现白化株对人离体皮肤组织的侵袭力较色素株下降,这表明黑色素能够增强皮炎外瓶霉的侵袭力(Poyntner et al. 2018).在真菌与细菌的研究中,真菌与细菌通过次级代谢产物相互作用中的例子有很多,细菌青枯雷尔氏菌Ralstonia solanacearum分泌脂肽ralsolamycin,其诱导真菌中的厚垣孢子形成和镰刀菌属中的bikaverin基因簇的表达,进而可减少细菌进入和生长(Spraker et al. 2016);研究发现伊氏杀线真菌胞内存在内生细菌,其合成的次级代谢产物将有助于伊氏杀线真菌在宿主线虫内定殖,增加伊氏杀线菌在生态竞争中的适应性,以便更好地捕杀线虫,减少松树枯萎病(王瑞珍 2017).在防御昆虫的过程中,真菌往往产生一些有毒的次级代谢产物来保证自身的安全.例如:白僵菌Beauveria bassiana可以通过其次级代谢产物白僵菌素杀死害虫,而且还用聚酮类卵磷脂毒化尸体,以限制其他微生物对其食物的竞争(Fan et al. 2017);同样的,黄曲霉产生的有毒次级代谢产物黄曲霉毒素对昆虫也有一定的杀伤力,研究发现,在黄曲霉毒素丰富的环境中,在与昆虫竞争的试验中,真菌的适应性提高了26倍,而黄曲霉产毒菌株显示出更高的适应性(Keller 2019). ...
Temperature during conidiation affects stress tolerance, pigmentation, and trypacidin accumulation in the conidia of the airborne pathogen Aspergillus fumigatus
1
2017
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Redox metabolites signal polymicrobial biofilm development via the NapA oxidative stress cascade in Aspergillus
1
2015
... 真菌产生的次级代谢产物与真菌的发育有着密切联系,上文中已经提到真菌黑色素不仅能够抵抗紫外线辐射,而且能够结合和隔离非特异性肽和化合物,产生保护细胞的物理屏障,为真菌孢子提供保护,使其免受极端环境或昆虫捕食.除此以外,研究表明,土曲霉产生的丁内酯,能在真菌中提高菌丝的分枝、孢子形成(Schimmel et al. 1998);在构巢曲霉中,内源性油酸和亚麻油酸衍生物能调控形成无性和有性孢子的比例(Calvo et al. 2001);镰刀菌属的色素形成需要镰刀菌素(Lena et al. 2012);神经孢菌素neurosporin A可以阻止食虫动物以粗糙脉孢菌孢子为食(Zhao et al. 2017).事实上,细菌的次级代谢产物也会对真菌发育产生影响,次级代谢产物可以作为种间信号,通过氧化应激调节影响真菌发育.在共培养实验中,铜绿假单胞菌次级代谢产物吩嗪对烟曲霉发育有不同的调节作用,其中高浓度的吩嗪具有抗真菌作用,但中等浓度会诱导真菌产孢(He et al. 2015). ...
TRI6 and TRI10 play different roles in the regulation of deoxynivalenol (DON) production by cAMP signalling in Fusarium graminearum
1
2016
... 此外,我们的前期研究和其他真菌中也发现真菌的次级代谢产物的合成受到了cAMP/PKA通路的调控作用(Yang et al. 2016;Yang et al. 2017).在构巢曲霉中,cAMP通路上游的Gα亚基FadA能够激活PKA,而PKA能够磷酸化AflR从而抑制它的活性,进而抑制ST/AF合成通路的开启,因此cAMP通路负调控ST/AF合成(Shimizu & Keller 2001;Roze et al. 2004).我们前期发现将黄曲霉菌中的cAMP高亲和力的磷酸二酯酶pdeH进行缺失后,导致黄曲霉内源cAMP浓度上升,但是由于黄曲霉菌中存在cAMP的高浓度反馈抑制作用导致突变体中的PKA酶活降低,并造成黄曲霉毒素产量都显著上升(Yang et al. 2017).禾谷镰刀菌中的研究表明,外源cAMP处理或者敲除pde2都能够提高菌内的PKA酶活,从而促进DON毒素合成基因TRI的转录,进而提高DON毒素的合成(Jiang et al. 2016).近期的研究也发现,蛋白的磷酸化、乙酰化、琥珀酰化和sumo化等翻译后修饰都参与了黄曲霉生长发育和AF合成过程(Nie et al. 2016;Ren et al. 2016;Lv 2017;Ren et al. 2018). ...
LaeA, a regulator of secondary metabolism in Aspergillus spp
1
2004
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Fungal secondary metabolism: regulation, function and drug discovery
4
2019
... 真菌在环境中无处不在,在植物、动物和人类的健康以及广泛的生态系统功能中都发挥着重要作用(Xu 2016).它们能产生各种各样的次级代谢产物,而次级代谢产物对其自身的正常生长不是必需的化合物,但其在真菌生态学中作为适应因子扮演着重要的角色,例如紫外线防护,并且能够通过在生态相互作用中作为防御化合物或信号分子起作用(Brakhage 2013;Keller 2019).不仅如此,真菌产生的这些次级代谢产物在人类生活中具有重要的应用,自弗莱明从青霉属Penicillium中发现青霉素并广泛应用以来,真菌次级代谢产物成为药物分子的重要来源(Raja et al. 2017),例如,他汀类、环孢菌素和霉酚酸等已经开始大规模地用于延长人类生命(Keller 2019).此外,真菌产生的次级代谢产物也表现出对人有害的特性,例如,黄曲霉毒素(AF)、环匹阿尼酸和黄曲霉震颤素等.由于次级代谢产物的重要性,所以对次级代谢产物的合成调控研究已经成为热点. ...
... ),例如,他汀类、环孢菌素和霉酚酸等已经开始大规模地用于延长人类生命(Keller 2019).此外,真菌产生的次级代谢产物也表现出对人有害的特性,例如,黄曲霉毒素(AF)、环匹阿尼酸和黄曲霉震颤素等.由于次级代谢产物的重要性,所以对次级代谢产物的合成调控研究已经成为热点. ...
... 次级代谢产物有利于真菌在激烈的竞争环境中生存,已经有大量研究表明真菌次级代谢产物在真菌与其他微生物、真菌与昆虫以及真菌与植物之间都起到非常重要的作用(Rohlfs 2015;Zeilinger et al. 2016;Scherlach & Hertweck 2017).如上文提到的次级代谢产物黑色素,不仅可以保护真菌孢子免受紫外线的伤害,而且可以作为毒力因子.有研究表明通过各种机制影响巨噬细胞在抗真菌免疫过程中的识别和吞噬等环节,降低其抗真菌效应,在烟曲霉感染中,黑色素可通过覆盖细胞壁表面的β-甘露糖,影响巨噬细胞PRRs对烟曲霉识别,帮助烟曲霉在巨噬细胞内存活(Akoumianaki et al. 2016);通过CRISPR/Cas9敲除皮炎外瓶霉的黑色素合成关键酶PKS1基因,也发现白化株对人离体皮肤组织的侵袭力较色素株下降,这表明黑色素能够增强皮炎外瓶霉的侵袭力(Poyntner et al. 2018).在真菌与细菌的研究中,真菌与细菌通过次级代谢产物相互作用中的例子有很多,细菌青枯雷尔氏菌Ralstonia solanacearum分泌脂肽ralsolamycin,其诱导真菌中的厚垣孢子形成和镰刀菌属中的bikaverin基因簇的表达,进而可减少细菌进入和生长(Spraker et al. 2016);研究发现伊氏杀线真菌胞内存在内生细菌,其合成的次级代谢产物将有助于伊氏杀线真菌在宿主线虫内定殖,增加伊氏杀线菌在生态竞争中的适应性,以便更好地捕杀线虫,减少松树枯萎病(王瑞珍 2017).在防御昆虫的过程中,真菌往往产生一些有毒的次级代谢产物来保证自身的安全.例如:白僵菌Beauveria bassiana可以通过其次级代谢产物白僵菌素杀死害虫,而且还用聚酮类卵磷脂毒化尸体,以限制其他微生物对其食物的竞争(Fan et al. 2017);同样的,黄曲霉产生的有毒次级代谢产物黄曲霉毒素对昆虫也有一定的杀伤力,研究发现,在黄曲霉毒素丰富的环境中,在与昆虫竞争的试验中,真菌的适应性提高了26倍,而黄曲霉产毒菌株显示出更高的适应性(Keller 2019). ...
... 真菌的次级代谢产物按照其合成途径主要分为聚酮类(polyketide)、非核糖体肽类(nonribosomal peptide)、生物碱以及萜类化合物等,次级代谢产物的合成主要通过骨干酶(backbone enzyme)聚合初级代谢产物,而骨干酶产生的代谢物会被其他能进一步改变代谢物生物活性的酶进一步“修饰”,骨干酶定义了产生的次级代谢产物的化学类别.例如,聚酮化合物合酶(polyketide synthases,PKS)从酰基CoA产生聚酮化合物,非核糖体肽合成酶(nonribosomal peptide synthases,NRPS)从氨基酸和萜烯合酶产生非核糖体肽,而萜烯环化酶(分别为TS和TC)从活化的异戊二烯单元产生萜烯(Keller 2019).总之,真菌次级代谢产物的合成是一个复杂的过程,具体涉及到真菌感受细胞外界环境信号,转录激活和翻译表达水平等,其中每一步都需要多个基因参与调控.参与真菌次级代谢物生物合成的基因通常排列在生物合成基因簇中,生物合成基因簇是一种含特定遗传信息的核苷酸序列,而且其结构基因受表观遗传调控. ...
Studies on secondary metabolites and their activities of endophytic fungi from Nerium indicum Mill. cv Paihua. PhD Dissertation, Shaanxi University of Science and Technology,
0
2019
LaeA regulation of secondary metabolism modulates virulence in Penicillium expansum and is mediated by sucrose
1
2017
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
The Aspergillus flavus histone acetyltransferase AflGcnE regulates morphogenesis, aflatoxin biosynthesis, and pathogenicity
1
2016
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Biosynthesis of fusarubins accounts for pigmentation of Fusarium fujikuroi perithecia
1
2012
... 真菌产生的次级代谢产物与真菌的发育有着密切联系,上文中已经提到真菌黑色素不仅能够抵抗紫外线辐射,而且能够结合和隔离非特异性肽和化合物,产生保护细胞的物理屏障,为真菌孢子提供保护,使其免受极端环境或昆虫捕食.除此以外,研究表明,土曲霉产生的丁内酯,能在真菌中提高菌丝的分枝、孢子形成(Schimmel et al. 1998);在构巢曲霉中,内源性油酸和亚麻油酸衍生物能调控形成无性和有性孢子的比例(Calvo et al. 2001);镰刀菌属的色素形成需要镰刀菌素(Lena et al. 2012);神经孢菌素neurosporin A可以阻止食虫动物以粗糙脉孢菌孢子为食(Zhao et al. 2017).事实上,细菌的次级代谢产物也会对真菌发育产生影响,次级代谢产物可以作为种间信号,通过氧化应激调节影响真菌发育.在共培养实验中,铜绿假单胞菌次级代谢产物吩嗪对烟曲霉发育有不同的调节作用,其中高浓度的吩嗪具有抗真菌作用,但中等浓度会诱导真菌产孢(He et al. 2015). ...
Isolation of secondary metabolites from macrofungi Hericium alpestre and anti-tumor effects and mechanism of 4-hydroxy-2-pyridone alkaloid sambutoxin. PhD Dissertation,
0
2019
5-Azacytidine inhibits aflatoxin biosynthesis in Aspergillus flavus
1
2013
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Bisulfite sequencing reveals that Aspergillus flavus holds a hollow in DNA methylation
1
2012
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
The velvet complex governs mycotoxin production and virulence of Fusarium oxysporum on plant and mammalian hosts
1
2013
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Proteome-wide profiling of protein lysine acetylation in Aspergillus flavus
1
2017
... 此外,我们的前期研究和其他真菌中也发现真菌的次级代谢产物的合成受到了cAMP/PKA通路的调控作用(Yang et al. 2016;Yang et al. 2017).在构巢曲霉中,cAMP通路上游的Gα亚基FadA能够激活PKA,而PKA能够磷酸化AflR从而抑制它的活性,进而抑制ST/AF合成通路的开启,因此cAMP通路负调控ST/AF合成(Shimizu & Keller 2001;Roze et al. 2004).我们前期发现将黄曲霉菌中的cAMP高亲和力的磷酸二酯酶pdeH进行缺失后,导致黄曲霉内源cAMP浓度上升,但是由于黄曲霉菌中存在cAMP的高浓度反馈抑制作用导致突变体中的PKA酶活降低,并造成黄曲霉毒素产量都显著上升(Yang et al. 2017).禾谷镰刀菌中的研究表明,外源cAMP处理或者敲除pde2都能够提高菌内的PKA酶活,从而促进DON毒素合成基因TRI的转录,进而提高DON毒素的合成(Jiang et al. 2016).近期的研究也发现,蛋白的磷酸化、乙酰化、琥珀酰化和sumo化等翻译后修饰都参与了黄曲霉生长发育和AF合成过程(Nie et al. 2016;Ren et al. 2016;Lv 2017;Ren et al. 2018). ...
Key role of LaeA and velvet complex proteins on expression of β-lactam and PR-toxin genes in Penicillium chrysogenum: cross-talk regulation of secondary metabolite pathways
1
2016
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
A non-linear model for temperature-dependent sporulation and T-2 and HT-2 production of Fusarium langsethiae and Fusarium sporotrichioides
1
2016
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Aspergillus flavus SUMO contributes to fungal virulence and toxin attributes
1
2016
... 此外,我们的前期研究和其他真菌中也发现真菌的次级代谢产物的合成受到了cAMP/PKA通路的调控作用(Yang et al. 2016;Yang et al. 2017).在构巢曲霉中,cAMP通路上游的Gα亚基FadA能够激活PKA,而PKA能够磷酸化AflR从而抑制它的活性,进而抑制ST/AF合成通路的开启,因此cAMP通路负调控ST/AF合成(Shimizu & Keller 2001;Roze et al. 2004).我们前期发现将黄曲霉菌中的cAMP高亲和力的磷酸二酯酶pdeH进行缺失后,导致黄曲霉内源cAMP浓度上升,但是由于黄曲霉菌中存在cAMP的高浓度反馈抑制作用导致突变体中的PKA酶活降低,并造成黄曲霉毒素产量都显著上升(Yang et al. 2017).禾谷镰刀菌中的研究表明,外源cAMP处理或者敲除pde2都能够提高菌内的PKA酶活,从而促进DON毒素合成基因TRI的转录,进而提高DON毒素的合成(Jiang et al. 2016).近期的研究也发现,蛋白的磷酸化、乙酰化、琥珀酰化和sumo化等翻译后修饰都参与了黄曲霉生长发育和AF合成过程(Nie et al. 2016;Ren et al. 2016;Lv 2017;Ren et al. 2018). ...
Secondary metabolic gene clusters: evolutionary toolkits for chemical innovation
1
2010
... 次级代谢产物的合成是一个复杂过程,需要多个基因参与调控,基因组挖掘工作表明,真菌产生次级代谢产物的能力被大大低估,因为许多真菌次级代谢产物生物合成基因簇在标准培养条件下是沉默的(Brakhage 2013).类似于原核生物的操纵子,在真菌的基因组中也出现了彼此功能相关的非同源基因成簇存在的现象.这些基因形成基因簇,可参与多种次级代谢途径(Osbourn 2010).真菌次级代谢基因簇由复杂的调节网络控制,其全局调控主要通过全局调控因子实现,而通路特异性调节由它们调节簇内的基因编码的转录因子介导.本文主要根据近年来对次级代谢产物的研究,综合论述了真菌次级代谢产物的药用价值以及在竞争、防御和发育过程中的生态功能,此外,还描述了次级代谢物生物合成基因簇的转录和表观遗传调控,旨在更加深刻地理解次级代谢产物的合成调控. ...
VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism
1
2008
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Transcriptome study of an Exophiala dermatitidis PKS1 mutant on an ex vivo skin model: is melanin important for infection?
1
2018
... 次级代谢产物有利于真菌在激烈的竞争环境中生存,已经有大量研究表明真菌次级代谢产物在真菌与其他微生物、真菌与昆虫以及真菌与植物之间都起到非常重要的作用(Rohlfs 2015;Zeilinger et al. 2016;Scherlach & Hertweck 2017).如上文提到的次级代谢产物黑色素,不仅可以保护真菌孢子免受紫外线的伤害,而且可以作为毒力因子.有研究表明通过各种机制影响巨噬细胞在抗真菌免疫过程中的识别和吞噬等环节,降低其抗真菌效应,在烟曲霉感染中,黑色素可通过覆盖细胞壁表面的β-甘露糖,影响巨噬细胞PRRs对烟曲霉识别,帮助烟曲霉在巨噬细胞内存活(Akoumianaki et al. 2016);通过CRISPR/Cas9敲除皮炎外瓶霉的黑色素合成关键酶PKS1基因,也发现白化株对人离体皮肤组织的侵袭力较色素株下降,这表明黑色素能够增强皮炎外瓶霉的侵袭力(Poyntner et al. 2018).在真菌与细菌的研究中,真菌与细菌通过次级代谢产物相互作用中的例子有很多,细菌青枯雷尔氏菌Ralstonia solanacearum分泌脂肽ralsolamycin,其诱导真菌中的厚垣孢子形成和镰刀菌属中的bikaverin基因簇的表达,进而可减少细菌进入和生长(Spraker et al. 2016);研究发现伊氏杀线真菌胞内存在内生细菌,其合成的次级代谢产物将有助于伊氏杀线真菌在宿主线虫内定殖,增加伊氏杀线菌在生态竞争中的适应性,以便更好地捕杀线虫,减少松树枯萎病(王瑞珍 2017).在防御昆虫的过程中,真菌往往产生一些有毒的次级代谢产物来保证自身的安全.例如:白僵菌Beauveria bassiana可以通过其次级代谢产物白僵菌素杀死害虫,而且还用聚酮类卵磷脂毒化尸体,以限制其他微生物对其食物的竞争(Fan et al. 2017);同样的,黄曲霉产生的有毒次级代谢产物黄曲霉毒素对昆虫也有一定的杀伤力,研究发现,在黄曲霉毒素丰富的环境中,在与昆虫竞争的试验中,真菌的适应性提高了26倍,而黄曲霉产毒菌株显示出更高的适应性(Keller 2019). ...
Fungal identification using molecular tools: a primer for the natural products research community
1
2017
... 真菌在环境中无处不在,在植物、动物和人类的健康以及广泛的生态系统功能中都发挥着重要作用(Xu 2016).它们能产生各种各样的次级代谢产物,而次级代谢产物对其自身的正常生长不是必需的化合物,但其在真菌生态学中作为适应因子扮演着重要的角色,例如紫外线防护,并且能够通过在生态相互作用中作为防御化合物或信号分子起作用(Brakhage 2013;Keller 2019).不仅如此,真菌产生的这些次级代谢产物在人类生活中具有重要的应用,自弗莱明从青霉属Penicillium中发现青霉素并广泛应用以来,真菌次级代谢产物成为药物分子的重要来源(Raja et al. 2017),例如,他汀类、环孢菌素和霉酚酸等已经开始大规模地用于延长人类生命(Keller 2019).此外,真菌产生的次级代谢产物也表现出对人有害的特性,例如,黄曲霉毒素(AF)、环匹阿尼酸和黄曲霉震颤素等.由于次级代谢产物的重要性,所以对次级代谢产物的合成调控研究已经成为热点. ...
Global phosphoproteomic analysis reveals the involvement of phosphorylation in aflatoxins biosynthesis in the pathogenic fungus Aspergillus flavus
1
2016
... 此外,我们的前期研究和其他真菌中也发现真菌的次级代谢产物的合成受到了cAMP/PKA通路的调控作用(Yang et al. 2016;Yang et al. 2017).在构巢曲霉中,cAMP通路上游的Gα亚基FadA能够激活PKA,而PKA能够磷酸化AflR从而抑制它的活性,进而抑制ST/AF合成通路的开启,因此cAMP通路负调控ST/AF合成(Shimizu & Keller 2001;Roze et al. 2004).我们前期发现将黄曲霉菌中的cAMP高亲和力的磷酸二酯酶pdeH进行缺失后,导致黄曲霉内源cAMP浓度上升,但是由于黄曲霉菌中存在cAMP的高浓度反馈抑制作用导致突变体中的PKA酶活降低,并造成黄曲霉毒素产量都显著上升(Yang et al. 2017).禾谷镰刀菌中的研究表明,外源cAMP处理或者敲除pde2都能够提高菌内的PKA酶活,从而促进DON毒素合成基因TRI的转录,进而提高DON毒素的合成(Jiang et al. 2016).近期的研究也发现,蛋白的磷酸化、乙酰化、琥珀酰化和sumo化等翻译后修饰都参与了黄曲霉生长发育和AF合成过程(Nie et al. 2016;Ren et al. 2016;Lv 2017;Ren et al. 2018). ...
Lysine succinylation contributes to aflatoxin 6 production and pathogenicity in Aspergillus flavus
1
2018
... 此外,我们的前期研究和其他真菌中也发现真菌的次级代谢产物的合成受到了cAMP/PKA通路的调控作用(Yang et al. 2016;Yang et al. 2017).在构巢曲霉中,cAMP通路上游的Gα亚基FadA能够激活PKA,而PKA能够磷酸化AflR从而抑制它的活性,进而抑制ST/AF合成通路的开启,因此cAMP通路负调控ST/AF合成(Shimizu & Keller 2001;Roze et al. 2004).我们前期发现将黄曲霉菌中的cAMP高亲和力的磷酸二酯酶pdeH进行缺失后,导致黄曲霉内源cAMP浓度上升,但是由于黄曲霉菌中存在cAMP的高浓度反馈抑制作用导致突变体中的PKA酶活降低,并造成黄曲霉毒素产量都显著上升(Yang et al. 2017).禾谷镰刀菌中的研究表明,外源cAMP处理或者敲除pde2都能够提高菌内的PKA酶活,从而促进DON毒素合成基因TRI的转录,进而提高DON毒素的合成(Jiang et al. 2016).近期的研究也发现,蛋白的磷酸化、乙酰化、琥珀酰化和sumo化等翻译后修饰都参与了黄曲霉生长发育和AF合成过程(Nie et al. 2016;Ren et al. 2016;Lv 2017;Ren et al. 2018). ...
Fungal secondary metabolite dynamics in fungus-grazer interactions: novel insights and unanswered questions
1
2015
... 次级代谢产物有利于真菌在激烈的竞争环境中生存,已经有大量研究表明真菌次级代谢产物在真菌与其他微生物、真菌与昆虫以及真菌与植物之间都起到非常重要的作用(Rohlfs 2015;Zeilinger et al. 2016;Scherlach & Hertweck 2017).如上文提到的次级代谢产物黑色素,不仅可以保护真菌孢子免受紫外线的伤害,而且可以作为毒力因子.有研究表明通过各种机制影响巨噬细胞在抗真菌免疫过程中的识别和吞噬等环节,降低其抗真菌效应,在烟曲霉感染中,黑色素可通过覆盖细胞壁表面的β-甘露糖,影响巨噬细胞PRRs对烟曲霉识别,帮助烟曲霉在巨噬细胞内存活(Akoumianaki et al. 2016);通过CRISPR/Cas9敲除皮炎外瓶霉的黑色素合成关键酶PKS1基因,也发现白化株对人离体皮肤组织的侵袭力较色素株下降,这表明黑色素能够增强皮炎外瓶霉的侵袭力(Poyntner et al. 2018).在真菌与细菌的研究中,真菌与细菌通过次级代谢产物相互作用中的例子有很多,细菌青枯雷尔氏菌Ralstonia solanacearum分泌脂肽ralsolamycin,其诱导真菌中的厚垣孢子形成和镰刀菌属中的bikaverin基因簇的表达,进而可减少细菌进入和生长(Spraker et al. 2016);研究发现伊氏杀线真菌胞内存在内生细菌,其合成的次级代谢产物将有助于伊氏杀线真菌在宿主线虫内定殖,增加伊氏杀线菌在生态竞争中的适应性,以便更好地捕杀线虫,减少松树枯萎病(王瑞珍 2017).在防御昆虫的过程中,真菌往往产生一些有毒的次级代谢产物来保证自身的安全.例如:白僵菌Beauveria bassiana可以通过其次级代谢产物白僵菌素杀死害虫,而且还用聚酮类卵磷脂毒化尸体,以限制其他微生物对其食物的竞争(Fan et al. 2017);同样的,黄曲霉产生的有毒次级代谢产物黄曲霉毒素对昆虫也有一定的杀伤力,研究发现,在黄曲霉毒素丰富的环境中,在与昆虫竞争的试验中,真菌的适应性提高了26倍,而黄曲霉产毒菌株显示出更高的适应性(Keller 2019). ...
Regulation of aflatoxin synthesis by FadA/cAMP/protein kinase A signaling in Aspergillus parasiticus
1
2004
... 此外,我们的前期研究和其他真菌中也发现真菌的次级代谢产物的合成受到了cAMP/PKA通路的调控作用(Yang et al. 2016;Yang et al. 2017).在构巢曲霉中,cAMP通路上游的Gα亚基FadA能够激活PKA,而PKA能够磷酸化AflR从而抑制它的活性,进而抑制ST/AF合成通路的开启,因此cAMP通路负调控ST/AF合成(Shimizu & Keller 2001;Roze et al. 2004).我们前期发现将黄曲霉菌中的cAMP高亲和力的磷酸二酯酶pdeH进行缺失后,导致黄曲霉内源cAMP浓度上升,但是由于黄曲霉菌中存在cAMP的高浓度反馈抑制作用导致突变体中的PKA酶活降低,并造成黄曲霉毒素产量都显著上升(Yang et al. 2017).禾谷镰刀菌中的研究表明,外源cAMP处理或者敲除pde2都能够提高菌内的PKA酶活,从而促进DON毒素合成基因TRI的转录,进而提高DON毒素的合成(Jiang et al. 2016).近期的研究也发现,蛋白的磷酸化、乙酰化、琥珀酰化和sumo化等翻译后修饰都参与了黄曲霉生长发育和AF合成过程(Nie et al. 2016;Ren et al. 2016;Lv 2017;Ren et al. 2018). ...
Mediators of mutualistic microbe-microbe interactions
1
2017
... 次级代谢产物有利于真菌在激烈的竞争环境中生存,已经有大量研究表明真菌次级代谢产物在真菌与其他微生物、真菌与昆虫以及真菌与植物之间都起到非常重要的作用(Rohlfs 2015;Zeilinger et al. 2016;Scherlach & Hertweck 2017).如上文提到的次级代谢产物黑色素,不仅可以保护真菌孢子免受紫外线的伤害,而且可以作为毒力因子.有研究表明通过各种机制影响巨噬细胞在抗真菌免疫过程中的识别和吞噬等环节,降低其抗真菌效应,在烟曲霉感染中,黑色素可通过覆盖细胞壁表面的β-甘露糖,影响巨噬细胞PRRs对烟曲霉识别,帮助烟曲霉在巨噬细胞内存活(Akoumianaki et al. 2016);通过CRISPR/Cas9敲除皮炎外瓶霉的黑色素合成关键酶PKS1基因,也发现白化株对人离体皮肤组织的侵袭力较色素株下降,这表明黑色素能够增强皮炎外瓶霉的侵袭力(Poyntner et al. 2018).在真菌与细菌的研究中,真菌与细菌通过次级代谢产物相互作用中的例子有很多,细菌青枯雷尔氏菌Ralstonia solanacearum分泌脂肽ralsolamycin,其诱导真菌中的厚垣孢子形成和镰刀菌属中的bikaverin基因簇的表达,进而可减少细菌进入和生长(Spraker et al. 2016);研究发现伊氏杀线真菌胞内存在内生细菌,其合成的次级代谢产物将有助于伊氏杀线真菌在宿主线虫内定殖,增加伊氏杀线菌在生态竞争中的适应性,以便更好地捕杀线虫,减少松树枯萎病(王瑞珍 2017).在防御昆虫的过程中,真菌往往产生一些有毒的次级代谢产物来保证自身的安全.例如:白僵菌Beauveria bassiana可以通过其次级代谢产物白僵菌素杀死害虫,而且还用聚酮类卵磷脂毒化尸体,以限制其他微生物对其食物的竞争(Fan et al. 2017);同样的,黄曲霉产生的有毒次级代谢产物黄曲霉毒素对昆虫也有一定的杀伤力,研究发现,在黄曲霉毒素丰富的环境中,在与昆虫竞争的试验中,真菌的适应性提高了26倍,而黄曲霉产毒菌株显示出更高的适应性(Keller 2019). ...
Effect of butyrolactone I on the producing fungus, Aspergillus terreus
1
1998
... 真菌产生的次级代谢产物与真菌的发育有着密切联系,上文中已经提到真菌黑色素不仅能够抵抗紫外线辐射,而且能够结合和隔离非特异性肽和化合物,产生保护细胞的物理屏障,为真菌孢子提供保护,使其免受极端环境或昆虫捕食.除此以外,研究表明,土曲霉产生的丁内酯,能在真菌中提高菌丝的分枝、孢子形成(Schimmel et al. 1998);在构巢曲霉中,内源性油酸和亚麻油酸衍生物能调控形成无性和有性孢子的比例(Calvo et al. 2001);镰刀菌属的色素形成需要镰刀菌素(Lena et al. 2012);神经孢菌素neurosporin A可以阻止食虫动物以粗糙脉孢菌孢子为食(Zhao et al. 2017).事实上,细菌的次级代谢产物也会对真菌发育产生影响,次级代谢产物可以作为种间信号,通过氧化应激调节影响真菌发育.在共培养实验中,铜绿假单胞菌次级代谢产物吩嗪对烟曲霉发育有不同的调节作用,其中高浓度的吩嗪具有抗真菌作用,但中等浓度会诱导真菌产孢(He et al. 2015). ...
Genetic involvement of a cAMP-dependent protein kinase in a G protein signaling pathway regulating morphological and chemical transitions in Aspergillus nidulans
1
2001
... 此外,我们的前期研究和其他真菌中也发现真菌的次级代谢产物的合成受到了cAMP/PKA通路的调控作用(Yang et al. 2016;Yang et al. 2017).在构巢曲霉中,cAMP通路上游的Gα亚基FadA能够激活PKA,而PKA能够磷酸化AflR从而抑制它的活性,进而抑制ST/AF合成通路的开启,因此cAMP通路负调控ST/AF合成(Shimizu & Keller 2001;Roze et al. 2004).我们前期发现将黄曲霉菌中的cAMP高亲和力的磷酸二酯酶pdeH进行缺失后,导致黄曲霉内源cAMP浓度上升,但是由于黄曲霉菌中存在cAMP的高浓度反馈抑制作用导致突变体中的PKA酶活降低,并造成黄曲霉毒素产量都显著上升(Yang et al. 2017).禾谷镰刀菌中的研究表明,外源cAMP处理或者敲除pde2都能够提高菌内的PKA酶活,从而促进DON毒素合成基因TRI的转录,进而提高DON毒素的合成(Jiang et al. 2016).近期的研究也发现,蛋白的磷酸化、乙酰化、琥珀酰化和sumo化等翻译后修饰都参与了黄曲霉生长发育和AF合成过程(Nie et al. 2016;Ren et al. 2016;Lv 2017;Ren et al. 2018). ...
Histone deacetylase activity regulates chemical diversity in Aspergillus
1
2007
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Adaptive melanin response of the soil fungus Aspergillus niger to UV radiation stress at “Evolution Canyon”, Mount Carmel, Israel
1
2008
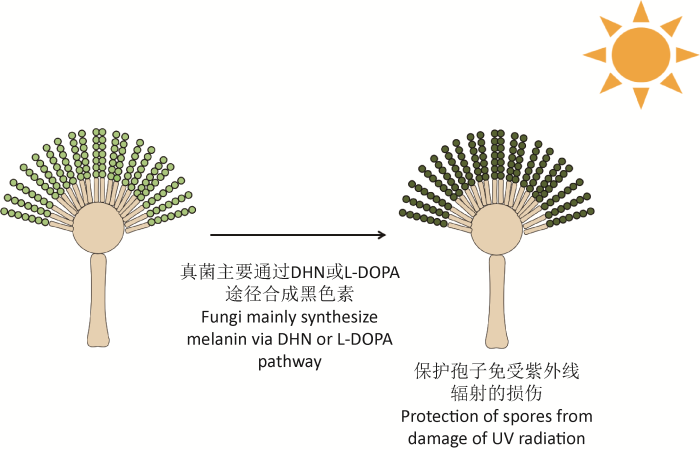
... 真菌黑色素是一种天然色素,一般呈黑棕色,通常在孢子或菌丝中发现,主要通过1,8-二羟基萘(DHN)途径和L-3,4-二羟基苯丙氨酸(L-DOPA)途径合成,还有一部分通过L-酪氨酸降解途径合成.研究表明,黑色素利于保护真菌孢子免受紫外线的伤害(图1),如玉米病菌Cochliobolus heterostrophus的白化突变体无法在田间存活;在黑曲霉中,破坏黑色素的合成会使其对紫外线的敏感度增加(Singaravelan et al. 2008).许多研究已经证明了真菌黑色素不仅能够抵抗紫外线辐射,而且能够结合和隔离非特异性肽和化合物,产生保护细胞的物理屏障. ...
Role of the Alternaria alternata blue-light receptor LreA (white-collar 1) in spore formation and secondary metabolism
1
2014
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Ralstonia solanacearum lipopeptide induces chlamydospore development in fungi and facilitates bacterial entry into fungal tissues
1
2016
... 次级代谢产物有利于真菌在激烈的竞争环境中生存,已经有大量研究表明真菌次级代谢产物在真菌与其他微生物、真菌与昆虫以及真菌与植物之间都起到非常重要的作用(Rohlfs 2015;Zeilinger et al. 2016;Scherlach & Hertweck 2017).如上文提到的次级代谢产物黑色素,不仅可以保护真菌孢子免受紫外线的伤害,而且可以作为毒力因子.有研究表明通过各种机制影响巨噬细胞在抗真菌免疫过程中的识别和吞噬等环节,降低其抗真菌效应,在烟曲霉感染中,黑色素可通过覆盖细胞壁表面的β-甘露糖,影响巨噬细胞PRRs对烟曲霉识别,帮助烟曲霉在巨噬细胞内存活(Akoumianaki et al. 2016);通过CRISPR/Cas9敲除皮炎外瓶霉的黑色素合成关键酶PKS1基因,也发现白化株对人离体皮肤组织的侵袭力较色素株下降,这表明黑色素能够增强皮炎外瓶霉的侵袭力(Poyntner et al. 2018).在真菌与细菌的研究中,真菌与细菌通过次级代谢产物相互作用中的例子有很多,细菌青枯雷尔氏菌Ralstonia solanacearum分泌脂肽ralsolamycin,其诱导真菌中的厚垣孢子形成和镰刀菌属中的bikaverin基因簇的表达,进而可减少细菌进入和生长(Spraker et al. 2016);研究发现伊氏杀线真菌胞内存在内生细菌,其合成的次级代谢产物将有助于伊氏杀线真菌在宿主线虫内定殖,增加伊氏杀线菌在生态竞争中的适应性,以便更好地捕杀线虫,减少松树枯萎病(王瑞珍 2017).在防御昆虫的过程中,真菌往往产生一些有毒的次级代谢产物来保证自身的安全.例如:白僵菌Beauveria bassiana可以通过其次级代谢产物白僵菌素杀死害虫,而且还用聚酮类卵磷脂毒化尸体,以限制其他微生物对其食物的竞争(Fan et al. 2017);同样的,黄曲霉产生的有毒次级代谢产物黄曲霉毒素对昆虫也有一定的杀伤力,研究发现,在黄曲霉毒素丰富的环境中,在与昆虫竞争的试验中,真菌的适应性提高了26倍,而黄曲霉产毒菌株显示出更高的适应性(Keller 2019). ...
Studies on comparative genomics and endobacteria of Esteva vermicola. PhD Dissertation,
0
2017
Resource diversity of Chinese macrofungi: edible, medicinal and poisonous species
1
2019
... 自古以来,人们就将真菌作为药材使用,《神农本草经》和《本草纲目》等书中就有记载,例如茯苓、灵芝等真菌药物.不仅如此,其产生的次级代谢产物在抗菌、抗心血管疾病、抗肿瘤和抗病毒等方面表现出不同的活性,具有重要的药用价值(Wu et al. 2019),其中一些次级代谢产物已经被开发成药物,例如:具有抗菌活性的药物青霉素、灰黄霉素就是从自然界中常见的青霉属Penicillium中分离得到,而卡泊芬净是由Glarea lozoyensis的发酵产物合成的;免疫抑制剂环孢菌素来自Trichoderma和Tolypocladium属;治疗心血管疾病的药物洛伐他汀类药物来自Aspergillus等属.还有许多次级代谢产物还没有被研发成药物,但是具有重要的药物活性,如陈旭等(2017)从追风伞植物中分离得一株木霉属内生真菌可以产甾体化合物,发现其能抑制两种癌细胞的增殖;孔阳(2019)从白花夹竹桃Nerium indicum mill. cv Paihua中分离出3株具有强抗菌活性的内生真菌,实验结果显示其次级代谢产物不仅对人前列腺癌细胞PC3、人肺癌细胞A549和人乳腺癌细胞MCF-7有良好的抑制效果,而且有一定的抗糖尿病活性;大型真菌Hericium alpestre产生的次级代谢产物——4-羟基-2-吡啶酮类生物碱sambutoxin具有良好的抗肿瘤作用(李璐宁 2019);周岳(2018)从海洋真菌Aspergillus versicolor LZD4403中分离得到的次级代谢产物曲林菌素可以通过抑制巨噬细胞炎症,进而发挥抑制巨噬细胞泡沫化作用,从而发挥抗动脉粥样硬化的作用;近年来研究发现真菌次级代谢产物黑色素在体外对艾滋病病毒有显著的抑制作用,这使天然黑色素有望成为一种新的抗艾滋病药物(Cordero & Casadevall 2017).以上研究均表明,真菌次级代谢产物在人类医疗健康方面具有重大价值,因此,了解真菌次级代谢产物的功能和合成调控具有重大意义. ...
Fungal DNA barcoding
1
2016
... 真菌在环境中无处不在,在植物、动物和人类的健康以及广泛的生态系统功能中都发挥着重要作用(Xu 2016).它们能产生各种各样的次级代谢产物,而次级代谢产物对其自身的正常生长不是必需的化合物,但其在真菌生态学中作为适应因子扮演着重要的角色,例如紫外线防护,并且能够通过在生态相互作用中作为防御化合物或信号分子起作用(Brakhage 2013;Keller 2019).不仅如此,真菌产生的这些次级代谢产物在人类生活中具有重要的应用,自弗莱明从青霉属Penicillium中发现青霉素并广泛应用以来,真菌次级代谢产物成为药物分子的重要来源(Raja et al. 2017),例如,他汀类、环孢菌素和霉酚酸等已经开始大规模地用于延长人类生命(Keller 2019).此外,真菌产生的次级代谢产物也表现出对人有害的特性,例如,黄曲霉毒素(AF)、环匹阿尼酸和黄曲霉震颤素等.由于次级代谢产物的重要性,所以对次级代谢产物的合成调控研究已经成为热点. ...
The high-affinity phosphodiesterase PdeH regulates development and aflatoxin biosynthesis in Aspergillus flavus
2
2017
... 此外,我们的前期研究和其他真菌中也发现真菌的次级代谢产物的合成受到了cAMP/PKA通路的调控作用(Yang et al. 2016;Yang et al. 2017).在构巢曲霉中,cAMP通路上游的Gα亚基FadA能够激活PKA,而PKA能够磷酸化AflR从而抑制它的活性,进而抑制ST/AF合成通路的开启,因此cAMP通路负调控ST/AF合成(Shimizu & Keller 2001;Roze et al. 2004).我们前期发现将黄曲霉菌中的cAMP高亲和力的磷酸二酯酶pdeH进行缺失后,导致黄曲霉内源cAMP浓度上升,但是由于黄曲霉菌中存在cAMP的高浓度反馈抑制作用导致突变体中的PKA酶活降低,并造成黄曲霉毒素产量都显著上升(Yang et al. 2017).禾谷镰刀菌中的研究表明,外源cAMP处理或者敲除pde2都能够提高菌内的PKA酶活,从而促进DON毒素合成基因TRI的转录,进而提高DON毒素的合成(Jiang et al. 2016).近期的研究也发现,蛋白的磷酸化、乙酰化、琥珀酰化和sumo化等翻译后修饰都参与了黄曲霉生长发育和AF合成过程(Nie et al. 2016;Ren et al. 2016;Lv 2017;Ren et al. 2018). ...
... 进行缺失后,导致黄曲霉内源cAMP浓度上升,但是由于黄曲霉菌中存在cAMP的高浓度反馈抑制作用导致突变体中的PKA酶活降低,并造成黄曲霉毒素产量都显著上升(Yang et al. 2017).禾谷镰刀菌中的研究表明,外源cAMP处理或者敲除pde2都能够提高菌内的PKA酶活,从而促进DON毒素合成基因TRI的转录,进而提高DON毒素的合成(Jiang et al. 2016).近期的研究也发现,蛋白的磷酸化、乙酰化、琥珀酰化和sumo化等翻译后修饰都参与了黄曲霉生长发育和AF合成过程(Nie et al. 2016;Ren et al. 2016;Lv 2017;Ren et al. 2018). ...
Cyclase-associated protein cap with multiple domains contributes to mycotoxin biosynthesis and fungal virulence in Aspergillus flavus
1
2019
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Adenylate cyclase AcyA regulates development, aflatoxin biosynthesis and fungal virulence in Aspergillus flavus
1
2016
... 此外,我们的前期研究和其他真菌中也发现真菌的次级代谢产物的合成受到了cAMP/PKA通路的调控作用(Yang et al. 2016;Yang et al. 2017).在构巢曲霉中,cAMP通路上游的Gα亚基FadA能够激活PKA,而PKA能够磷酸化AflR从而抑制它的活性,进而抑制ST/AF合成通路的开启,因此cAMP通路负调控ST/AF合成(Shimizu & Keller 2001;Roze et al. 2004).我们前期发现将黄曲霉菌中的cAMP高亲和力的磷酸二酯酶pdeH进行缺失后,导致黄曲霉内源cAMP浓度上升,但是由于黄曲霉菌中存在cAMP的高浓度反馈抑制作用导致突变体中的PKA酶活降低,并造成黄曲霉毒素产量都显著上升(Yang et al. 2017).禾谷镰刀菌中的研究表明,外源cAMP处理或者敲除pde2都能够提高菌内的PKA酶活,从而促进DON毒素合成基因TRI的转录,进而提高DON毒素的合成(Jiang et al. 2016).近期的研究也发现,蛋白的磷酸化、乙酰化、琥珀酰化和sumo化等翻译后修饰都参与了黄曲霉生长发育和AF合成过程(Nie et al. 2016;Ren et al. 2016;Lv 2017;Ren et al. 2018). ...
Friends or foes? Emerging insights from fungal interactions with plants
1
2016
... 次级代谢产物有利于真菌在激烈的竞争环境中生存,已经有大量研究表明真菌次级代谢产物在真菌与其他微生物、真菌与昆虫以及真菌与植物之间都起到非常重要的作用(Rohlfs 2015;Zeilinger et al. 2016;Scherlach & Hertweck 2017).如上文提到的次级代谢产物黑色素,不仅可以保护真菌孢子免受紫外线的伤害,而且可以作为毒力因子.有研究表明通过各种机制影响巨噬细胞在抗真菌免疫过程中的识别和吞噬等环节,降低其抗真菌效应,在烟曲霉感染中,黑色素可通过覆盖细胞壁表面的β-甘露糖,影响巨噬细胞PRRs对烟曲霉识别,帮助烟曲霉在巨噬细胞内存活(Akoumianaki et al. 2016);通过CRISPR/Cas9敲除皮炎外瓶霉的黑色素合成关键酶PKS1基因,也发现白化株对人离体皮肤组织的侵袭力较色素株下降,这表明黑色素能够增强皮炎外瓶霉的侵袭力(Poyntner et al. 2018).在真菌与细菌的研究中,真菌与细菌通过次级代谢产物相互作用中的例子有很多,细菌青枯雷尔氏菌Ralstonia solanacearum分泌脂肽ralsolamycin,其诱导真菌中的厚垣孢子形成和镰刀菌属中的bikaverin基因簇的表达,进而可减少细菌进入和生长(Spraker et al. 2016);研究发现伊氏杀线真菌胞内存在内生细菌,其合成的次级代谢产物将有助于伊氏杀线真菌在宿主线虫内定殖,增加伊氏杀线菌在生态竞争中的适应性,以便更好地捕杀线虫,减少松树枯萎病(王瑞珍 2017).在防御昆虫的过程中,真菌往往产生一些有毒的次级代谢产物来保证自身的安全.例如:白僵菌Beauveria bassiana可以通过其次级代谢产物白僵菌素杀死害虫,而且还用聚酮类卵磷脂毒化尸体,以限制其他微生物对其食物的竞争(Fan et al. 2017);同样的,黄曲霉产生的有毒次级代谢产物黄曲霉毒素对昆虫也有一定的杀伤力,研究发现,在黄曲霉毒素丰富的环境中,在与昆虫竞争的试验中,真菌的适应性提高了26倍,而黄曲霉产毒菌株显示出更高的适应性(Keller 2019). ...
Production of a fungal furocoumarin by a polyketide synthase gene cluster confers the chemo-resistance of Neurospora crassa to the predation by fungivorous arthropods
1
2017
... 真菌产生的次级代谢产物与真菌的发育有着密切联系,上文中已经提到真菌黑色素不仅能够抵抗紫外线辐射,而且能够结合和隔离非特异性肽和化合物,产生保护细胞的物理屏障,为真菌孢子提供保护,使其免受极端环境或昆虫捕食.除此以外,研究表明,土曲霉产生的丁内酯,能在真菌中提高菌丝的分枝、孢子形成(Schimmel et al. 1998);在构巢曲霉中,内源性油酸和亚麻油酸衍生物能调控形成无性和有性孢子的比例(Calvo et al. 2001);镰刀菌属的色素形成需要镰刀菌素(Lena et al. 2012);神经孢菌素neurosporin A可以阻止食虫动物以粗糙脉孢菌孢子为食(Zhao et al. 2017).事实上,细菌的次级代谢产物也会对真菌发育产生影响,次级代谢产物可以作为种间信号,通过氧化应激调节影响真菌发育.在共培养实验中,铜绿假单胞菌次级代谢产物吩嗪对烟曲霉发育有不同的调节作用,其中高浓度的吩嗪具有抗真菌作用,但中等浓度会诱导真菌产孢(He et al. 2015). ...
A cytosine methyltransferase ortholog dmtA is involved in the sensitivity of Aspergillus flavus to environmental stresses
1
2017
... 真菌次级代谢产物的合成与外界环境刺激有密不可分的联系,已经有研究表明温度和光可以诱导或抑制次级代谢产物的合成.以黄曲霉次级代谢产物黄曲霉毒素为例,黄曲霉毒素簇(图2A)由大约30个不同的基因组成,位于染色体3的端粒附近.在对黄曲霉毒素的研究过程中发现,不同温度培养下黄曲霉产毒能力有所不同(Yang et al. 2019).除此之外,烟曲霉的次级代谢产物毒素trypacidin和免疫调节剂endocrocin是温度依赖性的(Berthier et al. 2013;Hagiwara et al. 2017),而镰刀菌属中次级代谢产物萜烯T-2毒素的产生也是温度依赖性的(Nazari et al. 2016).事实上,研究表明光在真菌次级代谢物的合成过程中,也起到信号传导的作用.例如,黄曲霉毒素和杂色曲霉素相关的生物合成基因簇就受到白光抑制(Ozgür et al. 2008),光信号可以通过负调控全局转录因子LaeA来调控aflR,由aflR编码的Zn(II)2Cys6型转录因子可以调节黄曲霉毒素和杂色曲霉素合成基因的表达(图2B);此外,交链孢菌Alternaria alternata产生的真菌毒素交替醇和替代毒素则受白光刺激(Sonja et al. 2014).真菌生物合成基因的转录一般由转录调控因子控制,可以分为全局性调控因子和特异性转录因子.目前,全局调控最有影响力的转录复合物是Velvet复合物,由LaeA(或Lae1)、VeA(或Vel1)和VelB(或Vel2)组成,LaeA已被证明是曲霉菌和其他丝状真菌的次级代谢的全局性调控因子,考虑到Velvet复合物对次级代谢物表达的全局影响,通过构建laeA缺失株进行实验,在构巢曲霉中敲除laeA能抑制杂色曲霉素和青霉素的合成(Jin & Keller 2004);在黄曲霉中敲除laeA基因能抑制黄曲霉毒素的合成(Amaike & Keller 2009);在轮枝样镰刀菌中敲除laeA基因会抑制bikaverin和fusarin的生物合成(Butchko et al. 2012);在尖孢镰刀菌中敲除laeA基因能抑制白僵菌素的生物合成(López-Berges et al. 2013);敲除产黄青霉中laeA基因会抑制产黄青霉中青霉素和PR毒素的生物合成(Martín 2016);在Penicillium expansum中,敲除laeA基因会抑制棒曲霉素的生物合成(Kumar et al. 2017),结果表明LaeA可以调节杂色曲霉素、黄曲霉毒素、青霉素、bikaverin、fusarin和白僵菌素等次级代谢产物的合成;而敲除veA基因会使一些代谢产物如:黄曲霉毒素、环匹阿尼酸和黄曲霉震颤毒素的合成受到明显抑制(Duran et al. 2007).表观遗传对真菌次级代谢产物的生物合成也具有调控作用,DNA和组蛋白的甲基化与乙酰化修饰是目前所知的真菌主要的表观遗传调控形式.研究表明,编码组蛋白去乙酰化酶的hdaA的缺失导致转录激活并导致多个生物合成基因簇及其产物的表达增加(Shwab et al. 2007);Lan et al.(2016)研究发现黄曲霉中的组蛋白H3乙酰化酶GcnE参与了AF的合成及其合成基因的激活;中山大学贺竹梅课题组发现,DNA甲基转移酶抑制剂和DNA甲基转移酶基因的敲除能够抑制AF的合成,但是进一步研究发现黄曲霉菌中的DNA甲基化含量较低,其可能没有直接参与AF合成的调控(Liu et al. 2012;Lin et al. 2013;Zhi et al. 2017). ...
Mechanism of a marine fungal secondary metabolite: asperlin prevents atherosclerosis. Master Thesis,
0
2018
一株马比木植物内生真菌Trichoderma sp.抗癌活性研究
1
2017
... 自古以来,人们就将真菌作为药材使用,《神农本草经》和《本草纲目》等书中就有记载,例如茯苓、灵芝等真菌药物.不仅如此,其产生的次级代谢产物在抗菌、抗心血管疾病、抗肿瘤和抗病毒等方面表现出不同的活性,具有重要的药用价值(Wu et al. 2019),其中一些次级代谢产物已经被开发成药物,例如:具有抗菌活性的药物青霉素、灰黄霉素就是从自然界中常见的青霉属Penicillium中分离得到,而卡泊芬净是由Glarea lozoyensis的发酵产物合成的;免疫抑制剂环孢菌素来自Trichoderma和Tolypocladium属;治疗心血管疾病的药物洛伐他汀类药物来自Aspergillus等属.还有许多次级代谢产物还没有被研发成药物,但是具有重要的药物活性,如陈旭等(2017)从追风伞植物中分离得一株木霉属内生真菌可以产甾体化合物,发现其能抑制两种癌细胞的增殖;孔阳(2019)从白花夹竹桃Nerium indicum mill. cv Paihua中分离出3株具有强抗菌活性的内生真菌,实验结果显示其次级代谢产物不仅对人前列腺癌细胞PC3、人肺癌细胞A549和人乳腺癌细胞MCF-7有良好的抑制效果,而且有一定的抗糖尿病活性;大型真菌Hericium alpestre产生的次级代谢产物——4-羟基-2-吡啶酮类生物碱sambutoxin具有良好的抗肿瘤作用(李璐宁 2019);周岳(2018)从海洋真菌Aspergillus versicolor LZD4403中分离得到的次级代谢产物曲林菌素可以通过抑制巨噬细胞炎症,进而发挥抑制巨噬细胞泡沫化作用,从而发挥抗动脉粥样硬化的作用;近年来研究发现真菌次级代谢产物黑色素在体外对艾滋病病毒有显著的抑制作用,这使天然黑色素有望成为一种新的抗艾滋病药物(Cordero & Casadevall 2017).以上研究均表明,真菌次级代谢产物在人类医疗健康方面具有重大价值,因此,了解真菌次级代谢产物的功能和合成调控具有重大意义. ...
白花夹竹桃内生真菌次生代谢产物及其活性的研究
1
2019
... 自古以来,人们就将真菌作为药材使用,《神农本草经》和《本草纲目》等书中就有记载,例如茯苓、灵芝等真菌药物.不仅如此,其产生的次级代谢产物在抗菌、抗心血管疾病、抗肿瘤和抗病毒等方面表现出不同的活性,具有重要的药用价值(Wu et al. 2019),其中一些次级代谢产物已经被开发成药物,例如:具有抗菌活性的药物青霉素、灰黄霉素就是从自然界中常见的青霉属Penicillium中分离得到,而卡泊芬净是由Glarea lozoyensis的发酵产物合成的;免疫抑制剂环孢菌素来自Trichoderma和Tolypocladium属;治疗心血管疾病的药物洛伐他汀类药物来自Aspergillus等属.还有许多次级代谢产物还没有被研发成药物,但是具有重要的药物活性,如陈旭等(2017)从追风伞植物中分离得一株木霉属内生真菌可以产甾体化合物,发现其能抑制两种癌细胞的增殖;孔阳(2019)从白花夹竹桃Nerium indicum mill. cv Paihua中分离出3株具有强抗菌活性的内生真菌,实验结果显示其次级代谢产物不仅对人前列腺癌细胞PC3、人肺癌细胞A549和人乳腺癌细胞MCF-7有良好的抑制效果,而且有一定的抗糖尿病活性;大型真菌Hericium alpestre产生的次级代谢产物——4-羟基-2-吡啶酮类生物碱sambutoxin具有良好的抗肿瘤作用(李璐宁 2019);周岳(2018)从海洋真菌Aspergillus versicolor LZD4403中分离得到的次级代谢产物曲林菌素可以通过抑制巨噬细胞炎症,进而发挥抑制巨噬细胞泡沫化作用,从而发挥抗动脉粥样硬化的作用;近年来研究发现真菌次级代谢产物黑色素在体外对艾滋病病毒有显著的抑制作用,这使天然黑色素有望成为一种新的抗艾滋病药物(Cordero & Casadevall 2017).以上研究均表明,真菌次级代谢产物在人类医疗健康方面具有重大价值,因此,了解真菌次级代谢产物的功能和合成调控具有重大意义. ...
大型真菌Hericium alpestre次级代谢产物的分离和4-羟基-2-吡啶酮类生物碱sambutoxin的抗肿瘤作用及机制研究
1
2019
... 自古以来,人们就将真菌作为药材使用,《神农本草经》和《本草纲目》等书中就有记载,例如茯苓、灵芝等真菌药物.不仅如此,其产生的次级代谢产物在抗菌、抗心血管疾病、抗肿瘤和抗病毒等方面表现出不同的活性,具有重要的药用价值(Wu et al. 2019),其中一些次级代谢产物已经被开发成药物,例如:具有抗菌活性的药物青霉素、灰黄霉素就是从自然界中常见的青霉属Penicillium中分离得到,而卡泊芬净是由Glarea lozoyensis的发酵产物合成的;免疫抑制剂环孢菌素来自Trichoderma和Tolypocladium属;治疗心血管疾病的药物洛伐他汀类药物来自Aspergillus等属.还有许多次级代谢产物还没有被研发成药物,但是具有重要的药物活性,如陈旭等(2017)从追风伞植物中分离得一株木霉属内生真菌可以产甾体化合物,发现其能抑制两种癌细胞的增殖;孔阳(2019)从白花夹竹桃Nerium indicum mill. cv Paihua中分离出3株具有强抗菌活性的内生真菌,实验结果显示其次级代谢产物不仅对人前列腺癌细胞PC3、人肺癌细胞A549和人乳腺癌细胞MCF-7有良好的抑制效果,而且有一定的抗糖尿病活性;大型真菌Hericium alpestre产生的次级代谢产物——4-羟基-2-吡啶酮类生物碱sambutoxin具有良好的抗肿瘤作用(李璐宁 2019);周岳(2018)从海洋真菌Aspergillus versicolor LZD4403中分离得到的次级代谢产物曲林菌素可以通过抑制巨噬细胞炎症,进而发挥抑制巨噬细胞泡沫化作用,从而发挥抗动脉粥样硬化的作用;近年来研究发现真菌次级代谢产物黑色素在体外对艾滋病病毒有显著的抑制作用,这使天然黑色素有望成为一种新的抗艾滋病药物(Cordero & Casadevall 2017).以上研究均表明,真菌次级代谢产物在人类医疗健康方面具有重大价值,因此,了解真菌次级代谢产物的功能和合成调控具有重大意义. ...
伊氏线虫真菌比较基因组及其内生细菌的研究
1
2017
... 次级代谢产物有利于真菌在激烈的竞争环境中生存,已经有大量研究表明真菌次级代谢产物在真菌与其他微生物、真菌与昆虫以及真菌与植物之间都起到非常重要的作用(Rohlfs 2015;Zeilinger et al. 2016;Scherlach & Hertweck 2017).如上文提到的次级代谢产物黑色素,不仅可以保护真菌孢子免受紫外线的伤害,而且可以作为毒力因子.有研究表明通过各种机制影响巨噬细胞在抗真菌免疫过程中的识别和吞噬等环节,降低其抗真菌效应,在烟曲霉感染中,黑色素可通过覆盖细胞壁表面的β-甘露糖,影响巨噬细胞PRRs对烟曲霉识别,帮助烟曲霉在巨噬细胞内存活(Akoumianaki et al. 2016);通过CRISPR/Cas9敲除皮炎外瓶霉的黑色素合成关键酶PKS1基因,也发现白化株对人离体皮肤组织的侵袭力较色素株下降,这表明黑色素能够增强皮炎外瓶霉的侵袭力(Poyntner et al. 2018).在真菌与细菌的研究中,真菌与细菌通过次级代谢产物相互作用中的例子有很多,细菌青枯雷尔氏菌Ralstonia solanacearum分泌脂肽ralsolamycin,其诱导真菌中的厚垣孢子形成和镰刀菌属中的bikaverin基因簇的表达,进而可减少细菌进入和生长(Spraker et al. 2016);研究发现伊氏杀线真菌胞内存在内生细菌,其合成的次级代谢产物将有助于伊氏杀线真菌在宿主线虫内定殖,增加伊氏杀线菌在生态竞争中的适应性,以便更好地捕杀线虫,减少松树枯萎病(王瑞珍 2017).在防御昆虫的过程中,真菌往往产生一些有毒的次级代谢产物来保证自身的安全.例如:白僵菌Beauveria bassiana可以通过其次级代谢产物白僵菌素杀死害虫,而且还用聚酮类卵磷脂毒化尸体,以限制其他微生物对其食物的竞争(Fan et al. 2017);同样的,黄曲霉产生的有毒次级代谢产物黄曲霉毒素对昆虫也有一定的杀伤力,研究发现,在黄曲霉毒素丰富的环境中,在与昆虫竞争的试验中,真菌的适应性提高了26倍,而黄曲霉产毒菌株显示出更高的适应性(Keller 2019). ...
海洋真菌次级代谢产物曲林菌素抗动脉粥样硬化作用机制研究
1
2018
... 自古以来,人们就将真菌作为药材使用,《神农本草经》和《本草纲目》等书中就有记载,例如茯苓、灵芝等真菌药物.不仅如此,其产生的次级代谢产物在抗菌、抗心血管疾病、抗肿瘤和抗病毒等方面表现出不同的活性,具有重要的药用价值(Wu et al. 2019),其中一些次级代谢产物已经被开发成药物,例如:具有抗菌活性的药物青霉素、灰黄霉素就是从自然界中常见的青霉属Penicillium中分离得到,而卡泊芬净是由Glarea lozoyensis的发酵产物合成的;免疫抑制剂环孢菌素来自Trichoderma和Tolypocladium属;治疗心血管疾病的药物洛伐他汀类药物来自Aspergillus等属.还有许多次级代谢产物还没有被研发成药物,但是具有重要的药物活性,如陈旭等(2017)从追风伞植物中分离得一株木霉属内生真菌可以产甾体化合物,发现其能抑制两种癌细胞的增殖;孔阳(2019)从白花夹竹桃Nerium indicum mill. cv Paihua中分离出3株具有强抗菌活性的内生真菌,实验结果显示其次级代谢产物不仅对人前列腺癌细胞PC3、人肺癌细胞A549和人乳腺癌细胞MCF-7有良好的抑制效果,而且有一定的抗糖尿病活性;大型真菌Hericium alpestre产生的次级代谢产物——4-羟基-2-吡啶酮类生物碱sambutoxin具有良好的抗肿瘤作用(李璐宁 2019);周岳(2018)从海洋真菌Aspergillus versicolor LZD4403中分离得到的次级代谢产物曲林菌素可以通过抑制巨噬细胞炎症,进而发挥抑制巨噬细胞泡沫化作用,从而发挥抗动脉粥样硬化的作用;近年来研究发现真菌次级代谢产物黑色素在体外对艾滋病病毒有显著的抑制作用,这使天然黑色素有望成为一种新的抗艾滋病药物(Cordero & Casadevall 2017).以上研究均表明,真菌次级代谢产物在人类医疗健康方面具有重大价值,因此,了解真菌次级代谢产物的功能和合成调控具有重大意义. ...








{kind=link}
{kind=link}
{kind=link}
{kind=link}