



长期以来,我国重要药用真菌蝉花的分类地位及名称一直处于混乱状态。当前学界多认定蝉花是Miquel 1838年依据巴西标本命名的Isaria cicadae Miquel,对二者形态的差异颇欠考虑。因蝉花的有性型一直未发现,因而也影响对其分类地位的判断。过去或现在都有人将Cordyceps cicadae Massee(Massee 1895)视为I. cicadae的有性型而采用(Wu et al. 2019;Zha et al. 2019);因受日本文化影响,它曾被当作小蝉草Ophiocordyceps sobolifera的无性型而被称为Cordyceps sobolifera(蔡邦华 1957;刘波1974);它也曾被当作大蝉草(地方名“独角龙”)的无性型而采用Cordyceps cicadae Shing(=Tolypocladium dujiaolongae Y.P. Cao & C.R. Li)之名(幸兴球 1975;梁宗琦 2007);此外,还有人提出蝉花是小林虫草Cordyceps kobayasii Koval的无性型(Zha et al. 2018)。在GenBank中,虽有众多菌株的ITS序列及两个菌株的5基因序列,但材料皆为无性型。在Isaria属被证明是虫草科中的一个多系的类群(代永东等 2016)后,它在系统发育中已无独立位置。
我国学界寻找蝉花有性型多年,直到2015年7月13日才在江西井冈山发现。有些标本上同时存在有性型和无性型,无性型形态与我国南方各地采挖的药用蝉花一致,符合笔者以蝉棒束孢Isaria cicadae Miquel之名的详细描述(李增智等 2014),但与Miquel(1838)原描述的孢梗束极其简单的I. cicadae大相径庭。对井冈山标本及分离物的形态学、分类学和分子系统学研究,不仅有助于确定蝉花的分类地位,而且有助于辨明被认为世界广布的巴西种I. cicadae的多样性、复合性及在世界各地可能隐含的种类。
1 材料与方法
1.1 形态学研究
有性型描述依据寄主标本,无性型描述则根据纯培养物上的产孢结构。
1.1.1 供试标本:感染虫草菌的竹蝉Platylomia pieli Kato共7只。JGS150713号共5只,均在毛竹林中采集,其中1只(JGS150713-1)同时具有子座和孢梗束;JGS150725号,共2只,均同时具有子座和孢梗束。
1.1.2 供试菌种:BAIC1273号,从标本JGS150713-1子座分离。纯化后点植到Czapek-Dox和PSA培养基上,26℃下培养14d。
1.2 系统发育研究
1.2.1 DNA提取、ITS序列和5基因序列扩增及测序:有性型直接切取子座,采用无菌水冲洗后,采用CTAB法(刘少华等 2005)提取总DNA。无性型采用组织块分离法,切取部分孢梗束,接种至SDAY培养基,待菌丝长出后挑取菌落边缘菌丝纯化镜检后,接种至铺有玻璃纸的SDAY培养基上,培养5d后,刮取菌丝采用CTAB法提取基因组DNA。对ITS序列及LSU、SSU、RPB1、RPB2和TEF等5个基因片段进行PCR扩增和测序。参考White et al.(1990)的方法选择ITS4/ITS5用于扩增ITS;参考Vilgalys & Sun(1994)的方法选择LROR/LR5用于LSU、NS1和NS4用于扩增SSU;参考Castlebury et al.(2004)的方法选择RPB1A/RPB1C用于扩增RPB1;参考Rojas et al.(2010)的方法选择RPB2-5F/ RPB2-7cR用于扩增RPB2;参考Rehner & Buckley(2005)的方法选择983F/2218R用于扩增TEF。采用PCR Master Mix(Promega)的50μL体系,PCR反应条件参考相应文献进行,依据PCR结果对退火温度进行适当调整。扩增产物经0.8%琼脂糖凝胶电泳检测合格后送生工生物工程(上海)股份有限公司测序。
1.2.2 多位点系统发育分析:为确定井冈山标本的确切分类地位,参考Sung et al.(2007)、代永东等(2016)和Mongkolsamrita et al.(2018)的报道,从GenBank选取标为Cordyceps cicadae的4个菌株和1个标为Isaria cicadae的菌株的ITS位点和其他5基因位点序列,GenBank中唯一的Isaria sinclairii菌株的ITS序列,以及虫草属Cordyceps的其他22个分类单元的23个菌株(C. jakajanicola有2个菌株)的ITS位点和其他5基因位点序列进行多位点系统发育分析;为进一步澄清蝉花与曾被误认的小蝉草O. sobolifera和大蝉草(现称独角龙弯颈霉)T. dujiaolongae(幸辛球 1975;Li et al. 2018)的系统发育关系,也选取了它们及线虫草科冬虫夏草Ophiocordyceps sinensis各一个菌株的多位点序列。采用MUSCLE v3.8.31(Edgar 2004)对6个位点进行多序列比对,对每个位点的序列矩阵校对后,使用SequenceMatrix 1.8的方法(Vaidya et al. 2011)对ITS和5个基因序列进行拼接并导出序列矩阵。
数据集包括30个分类单元的38个菌株(含外群及井冈山材料),157条DNA序列的6位点矩阵,其中ITS 723bp、LSU 911bp、SSU 1 295bp、RPB1 698bp、RPB2 1 016bp和TEF 968bp;序列总长度达5 611bp(表1),1 776个独特模式,其中3 696个恒定点,1 518个简约信息点,397个单碱基差异点。使用IQ-TREE(Minh et al. 2020)软件包,采用最大似然法(Rehner & Buckley 2005),以肉座菌目丛赤壳科朱红丛赤壳Nectria cinnabarina作为外群,构建ML系统发育树。最大似然法用1 000次重复获得自举支持率,同时选用超快自展和1 000次SH-aLRT检验估算节点的可信度。经IQ-TREE计算,得到联合基因的贝叶斯信息准则(BIC)最适模型为:TIM3+F+G4。
表1 用于多位点系统发育分析的菌株
Table 1
| 物种 Taxon | 菌株号 Strain | 产地 Origin | GenBank检索号 GenBank accession number | |||||
|---|---|---|---|---|---|---|---|---|
| ITS | LSU | SSU | TEF | RPB1 | RPB2 | |||
| Beauveria bassiana | ARSEF 1478 | 巴西 Brazil | HQ880764 | HQ880836 | — | KJ523140 | KJ500427 | HQ880908 |
| Beauveria brongniartii | BCC 16585 | 泰国 Thailand | JN049867 | JF415967 | — | JN049885 | JF415991 | JF416009 |
| Cordyceps bifusispora | EFCC 8260 | 韩国 Korea | — | EF468807 | EF468953 | EF468855 | EF468910 | EF468747 |
| Cordyceps blackwelliae | TBRC 7253/ BBH23883 | 泰国 Thailand | MF140739 | MF140705 | — | MF140774 | MF140798 | MF140825 |
| Cordyceps cateniannulatus | TBRC 7258 | 泰国 Thailand | AY624173 | — | — | — | MG665236 | JQ425688 |
| Cordyceps cateniobliqua | CBS 153.83 T | 中国贵州 Guizhou, China | AY624173 | — | AY526466 | — | MG665236 | JQ425688 |
| Cordyceps cf. ochraceostromata | ARSEF 5691 | 韩国 Korea | — | EF468819 | EF468964 | EF468867 | EF468921 | EF468759 |
| Cordyceps chiangdaoensis | TBRC 7274 T | 泰国 Thailand | KT261393 | MF140732 | — | — | — | KT261403 |
| Cordyceps cicadae | ARSEF 7260 | 韩国 Korea | HQ880826 | — | — | HQ880898 | HQ880970 | HQ881017 |
| Cordyceps cicadae | GMSL 114 | 葡萄牙 Portugal | LT220698 | — | — | — | — | LT220790 |
| Cordyceps cicadae | YHICYY 101 | 中国云南 Yunnan, China | — | KP743160 | KP743168 | KP743140 | KP743147 | KP743152 |
| Cordyceps cicadae | RCEF HP090724 -31 | 中国广西 Guangxi, China | AF368801 | MF416552 | MF416605 | MF416496 | MF416653 | MF416447 |
| Cordyceps coleopterorum | CBS 110.73 T | 加纳 Ghana | AY624177 | JF415988 | — | JN049903 | JF416006 | JF416028 |
| Cordyceps farinosa | CBS 111113 T | 丹麦 Denmark | AY624181 | FJ765253 | — | — | GU979973 | GQ250022 |
| Cordyceps fumosorosea | CBS 244.31 | 爱尔兰 Ireland | AY624182 | MG665230 | MF416609 | — | — | JQ425690 |
| Cordyceps ghanensis | BS 105.73 T | 加纳 Ghana | AY624185 | — | MN296399 | — | — | — |
| Cordyceps jakajanicola | BCC79817 | 泰国 Thailand | — | MN275697 | MN296395 | MN338485 | MN338490 | MN338480 |
| Cordyceps jakajanicola | BCC79816 | 泰国 Thailand | — | MN275696 | MN296394 | MN338484 | MN338489 | MN338479 |
| Cordyceps javanica | TBRC 7260/ BBH 40412 | 泰国 Thailand | MF140744 | MF140710 | — | MF140779 | MF140803 | MF140830 |
| Cordyceps kintrischica | ARSEF 8058 | 格鲁吉亚 Georgia | GU734764 | — | — | — | — | GU734750 |
| Cordyceps kyusyuensis | EFCC 5886 | 韩国 Korea | — | EF468813 | EF468960 | EF468863 | EF468917 | EF468754 |
| Cordyceps militaris | OSC 93623 | 美国 USA | JN049825 | AY184966 | AY184977 | DQ522377 | — | DQ522332 |
| Cordyceps morakotii | TBRC 7276 T | 泰国 Thailand | KT261390 | MF140731 | — | — | — | KT261400 |
| Cordyceps ninchukispora | EGS 38.165 | 中国台湾 Taiwan, China | — | EF468846 | EF468991 | EF468900 | — | EF468795 |
| Cordyceps oncoperae | AFSEF 4358 | 澳大利亚 Australia | — | AF339532 | — | EF468891 | EU369083 | DQ118749 |
| Cordyceps piperis | CBS 116719 | 巴拿马 Panama | — | AY466442 | — | DQ127240 | EU369083 | DQ118749 |
| Cordyceps pruinosa | ARSEF 5413 | 中国贵州 Guizhou, China | JN049826 | AY184968 | AY184979 | DQ522397 | DQ522451 | DQ522351 |
| Cordyceps takaomontana | BCC 28612 | 泰国 Thailand | FJ765285 | FJ765252 | — | — | — | FJ765268 |
| Cordyceps tenuipes | ARSEF 5135 T | 美国 USA | AY624196 | JF415980 | — | JN049896 | JF416000 | JF416020 |
| Isaria cicadae | BA-001 | 中国安徽 Anhui, China | MT555325 | MT555409 | — | MT637810 | — | MT637807 |
| Isaria sinclairii | ATCC 24400 | ? | KU933437 | KU933411 | — | — | — | KU933431 |
| Nectria cinnabarina | G.J.S. 91-121 | 美国 USA | HM484697 | HM484753 | — | HM484770 | — | HM484660 |
| Ophiocordyceps sinensis | QH09 201 | 中国青海 Qinghai, China | JQ325080 | JX968029 | JX968024 | JX968004 | JX968009 | JX968014 |
| Ophiocordyceps sobolifera | NBRC 106967 | 中国安徽 Anhui, China | AB968409 | AB968422 | AB968395 | — | AB968551 | AB968590 |
| Tolypocladium dujiaolongae | YPC-2013 TEL | 中国安徽 Anhui, China | KF696557 | — | — | — | — | — |
| Tolypocladium dujiaolongae | YPC-2013 ANA | 中国安徽 Anhui, China | KF696558 | — | — | — | — | — |
| Cordyceps chanhua | JGS150713-1 TEL | 中国江西 Jiangxi, China | MT192488 | MT239107 | — | MT268246 | MT268242 | MT268244 |
| Cordyceps chanhua | JGS150713-1 ANA | 中国江西 Jiangxi, China | MT192487 | MT239106 | — | MT268245 | MT268241 | MT268243 |
2 结果与分析
2.1 多位点系统发育分析
30个分类单元的38个菌株的6位点系统发育分析所获得的ML系统树(图1)清楚地显示,井冈山的蝉花标本(JGS150713-1 TEL)和分离株(JGS150713-1 ANA)与浙江泛亚生物医药的蝉花生产菌株BA001以100%的支持率形成虫草科里的一个单独分支。BA001的全基因组序列已由该公司测定,并于2011年3月16日使用P. cicadae之名在香港发布。因此,这一分支的多位点序列和GenBank上的全基因组序列(检索号:PRJNA53245)典型地代表了我国古老的药用真菌蝉花的DNA特征。
图1
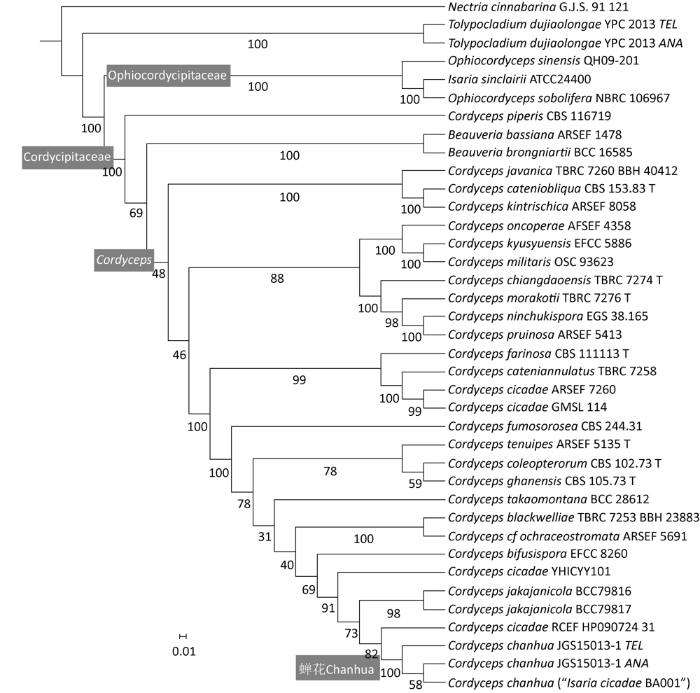
图1
蝉花有性型(JGS0713-1 TEL)和无性型(JGS0713-1 ANA)与虫草属其他物种的多位点系统发育关系
Fig. 1
Multilocal phylogenetic relationship of the teleomorph and anamorph of Cordyceps chanhua to other cordycipitoid species.
从系统树中可见,历史上曾被误认为是蝉花的小蝉草和大蝉草(独角龙弯颈霉)都不属于蝉花所在的虫草科。小蝉草属于线虫草科,而大蝉草(独角龙弯颈霉)则不仅不属于虫草科,而且还有可能不属于线虫草科,需要进一步研究确定。
在系统树中,蝉花分支与其他已见诸报道的C. cicadae菌株分开。与该分支最近的广西猫儿山菌株C. cicadae RCEF HP090724-31(Kepler et al. 2017)支持率为82%,与代永东等(2016)报道的云南菌株YHICYY 101的关系甚至远于子囊孢子形态截然不同的泰蝉虫草Cordyceps jakajanicola。蝉花与Sharma et al.(2018)报道的葡萄牙菌株GMSL 114和Rehner et al.(2011)报道的韩国菌株ARSEF 7260相距更远。这从系统发育上证明了本系列论文第1篇(李增智等 2020)里提出的质疑,即Samson(1974)组合的Paecilomyces cicadae和当代广为使用的Isaria (Paecilomyces) cicadae(Luangsa-ard et al. 2005)是一个物种复合群(species complex)。
2.2 形态描述
从子座上和孢梗束上分离出的无性型之间以及与BA001菌株之间的培养特征和显微特征一致。
蝉花 新种
Cordyceps chanhua Z.Z. Li, F.G. Luan, Hywel- Jones, C.R. Li & S.L. Zhang, sp. nov.
MycoBank MB835243
子座:从蝉若虫头部及前胸背板各长出一个具淡橙色或淡桔黄色的柄,多次二歧分枝,可形成淡桔黄色或橙色的可孕的头1-8个以上。也可二歧分枝同时产生子座和孢梗束。子囊壳半埋生,拟卵形,外壁单层,透明,长476-602μm,宽222-319μm,无间丝。子囊无色透明,圆筒形,有深裂的子囊帽,成熟时长235-380μm,宽2-3μm。子囊孢子8个,光滑,丝状,无色透明,长247-360μm,宽1.5-1.8μm,断裂成6.4-13.8×2.1-3.1μm长的分孢子。
孢梗束:出土前从蝉若虫头部及前胸背板长出1-3个柱状的柄(图2A),顶部尖削;有时也可从子座上长出;柄新鲜时淡黄色,干燥后深褐色。出土后延伸到一定长度后开始多次总状分枝,分枝柄部淡黄白色至橙黄色,干燥后淡黄褐色,长9-38mm,宽2-3.5mm,端部淡黄白色,形成由6-25个长30-80mm、宽2-6mm的分枝组成的鸡冠花状或西蓝花状的致密产孢结构,密布枯草黄色分生孢子。也可先在土壤中形成淡褐色的较粗壮的菌索,出土后再从菌索上长出孢梗束。在孢梗束形成过程中,寄主体表局部或通体覆盖有菌丝组成的菌膜。
图2

图2
蝉花的形态 A,B:长有完整子座和孢梗束的新鲜标本(A)和干标本(B);C:只有子座的标本;D,E:子座细部;F:半埋生的子囊壳;G:压片下的子囊壳呈埋生状;H:子囊;I-K:子囊孢子;J:子囊孢子断裂成分孢子;L,M:PSA培养基上的分生孢子梗和链生的分生孢子;N:标本上的分生孢子梗和链生的分生孢子;O:未成熟的厚垣孢子;P:成熟的厚垣孢子
Fig. 2
Morphology of Cordyceps chanhua. A, B: Fresh (A) and dry (B) specimens with both stromata and conidiophores; C: A specimen with only stromata; D, E: Stroma in detail; F: Subimmersed perithecia; G: Pseudo-immersed perithecia under coverslip; H: Asci; I-K: Ascospres; J: Part spores from broken ascospores; L, M: Conidiophores and conidia in chain on PSA medium; N: Conidiophores and conidia in chain from specimen; O: Immature chlamydospores; P: Mature chlamydospores.
菌落:24℃下在Czapek-Dox培养基上14d直径达49-55mm,在PSA培养基上14d直径达60-72mm,黄白色至淡黄色,绒毛状,具明显的轮纹或辐射纹,背面略呈白色;渗出液水珠状,无色;后期因产生大量分生孢子致使菌落表面呈粉状,淡黄白色。菌丝管状,分隔,壁光滑,无色透明,宽2.0-3.0μm。分生孢子梗轮状分枝为2-5个不规则的瓶梗(产孢细胞);标本上的瓶梗基部多为球形膨大,培养物上的瓶梗多为瓶形膨大,皆向上形成突然变细的颈部,偶呈锥状尖削;宽0.5μm,4.2-7(13.5)×2.3-3.5(5.2)μm。分生孢子形成向基型的链,单细胞,长椭圆形或圆筒形,两端钝圆,多对称,偶弯曲,3.5-10.5× 1.5-4.5μm,长径比2.4(2.2-2.8)。在老培养物上形成厚垣孢子,单生,棒状,肾形或茄形,单细胞,淡褐色,13-26.5×3-12μm,多光滑;成熟时椭球形或球形,暗褐色,直径6-18μm,有脐,具刺;附着在容器壁上聚集成由黑色小颗粒组成的环状物,易被误认为污染物。
模式标本:JGS150713-1,寄主:竹蝉Platylomia pieli Kato(同翅目:蝉科),采自江西省井冈山的毛竹林,东经114°07’,北纬26°34’,海拔961m。采集时间2015年7月13日,采集人:栾丰刚。模式标本及干培养物保存在浙江泛亚生命科学研究院虫生真菌标本室(ZBAH)。
名称词源:chanhua,蝉花的汉字拼音,古药材名,源自公元5世纪雷斅《雷公炮炙论》。
Cordyceps chanhua Z.Z. Li, F.G. Luan, Hywel-Jones, C.R. Li & S.L. Zhang, sp. nov.
Stromata solitary, simple from dead cicada or from rhizomorph, with fertile part orange to saffron, stipe light orange to light saffron, and base light yellowish, cylindric or flattened clavate. Perithecia partly immersed, ovoid with single-layered hyaline wall, 476-602×222-319μm. Hamathecia absent. Asci hyaline, cylindric, capitate, 8-spored, 235-380μm long and 2.1-3μm wide when mature, ascospores smooth, filiform, hyaline, 8-spored, 247-360μm long and 1.5-1.8μm wide, breaking into part spores, 6.4-13.8× 2.1-3.1μm.
Colonies attaining a diameter of 49-55mm on Czapek-Dox and 60-72mm on PDA after 14 days at 24°C, pale yellowish to yellowish, villous, with clear rings or radial lines. Reverse colorless. Synnemata pale yellowish, consisting of many erect or slightly curved branches emerging from front part of cadavers, especially from head; the branches cylindrical with tapering tip before emerging from soil and gradually branching into clusters of 6-25 branches after emerging, 30-80mm long, 2-6mm wide; stalks yellowish when fresh and dark brown after drying. Conidiophores consisting of verticillate branches with whorls of 2 to 5 phialides. Phialides 4.2-7 (-13.5)×2.3-3.5(-5.2)μm, consisting of a globose, oval or occasionally conical swollen basal portion tapering suddenly into a thin neck 0.5μm wide. Conidia in dry basipetal chains, one celled, long ellipsoid or cylindrical, mostly symmetrical, rarely curved, 4.2-7(13.5)× 2.3-3.5(5.2)μm. Chlamydospores solitary, clavate, cylindrical, reniform or egg plant shaped, unicellular, 13-26.5×3-12μm, with wall mostly smooth when immature, ellipsoidal or spherical, dark brown, 6-18μm in diam., and spinose with a navel when mature.
Holotype: JGS150713-1, on bamboo cicada Platylomia pieli, Kato (Hemiptera: Cicadidae, collected by Fenggang Luan, in a bamboo plantation in Mt Jinggangshan, 114°07’ E,26°34’ N,961m above sea level, 13-07-2015. Type specimen and dry culture stored at ZBAH, Zhejiang BioAsia Herbarium, Zhajiang BioAsia Pharmaceutical Co., Ltd.
Etymology: Chanhua, cicada flower in Chinese, an ancient Chinese medical fungus, originating in Leixiao’s Mr. Lei's Preparation Method, written in 5th century AD.
2.3 同近似种的比较
从子座宏观形态上看,蝉花与小林虫草Cordyceps kobayasii Koval(=C. sinclairii Kobayasi)相似,但其475-602×222-319μm的子囊壳比小林虫草的270-330×180-195μm大;而其235-380×2-3μm的子囊比小林虫草的150-200×8-13μm细长。显然,它们不是同一种虫草。
蝉花的DNA近似种泰蝉虫草子囊孢子极其独特,长达250-310μm,流星锤状,即中间形成直径仅1μm宽的细长的腰,而两端膨大成梭状;其分生孢子比蝉花小,稍细长。它们显然是不同的物种。
另外,刘爱英等(2011)曾发表“虫草属一新种,蝉棒束孢有性型”一文。根据刘爱英(2012)的描述,这个从广西乐业县神木天坑中采集的标本子囊壳676-718.5× 338-402μm,远大于蝉花,但无子囊孢子大小的量度,分孢子2-3×1-1.5μm,远小于蝉花。因此,这个神木蝉虫草新种Cordyceps cicadae-SM AY Liu & X Zou不是本文描述的蝉花有性型。
3 讨论
Miquel(1838)描述的I. cicadae的巴西模式标本极其简单,连分生孢子的量度都没有,插图也过于简单、模糊。而Samson(1974)在将其组合为Paecilomyces cicadae时,其详细形态描述及插图是根据对他所组合的其他物种标本的观察,因此不可避免地反映了混进的其他物种的性状,例如从其插图中即可发现,三排分生孢子中有两排是圆筒形,而第三排竟然都是弯曲的肾形!按照他宽广的描述,P. cicadae的分生孢子3.5-8.0× 1.5-3.5μm,从而与其他一些种多有重叠:Cunninghum(1921)描述新西兰的I. sinclairii的分生孢子长7-8μm;而Kobayasi(1939)从日本标本描述的I. sinclairii的分生孢子仅稍短而已,大小为5.9×2-3μm;与Koval描述远东标本的6-7×2-2.5μm间差别也不大。几个描述的分生孢子几乎都是圆筒形;而井冈山标本分离株也是圆筒形,大小为3.5-10.5× 1.5-4.5μm,因此,单从分生孢子的量度看是难以区别的。在传统的形态分类学时代,他把产自亚非拉的多个只了解无性型形态的近似种都处理为新组合可以理解;但在分子时代,通过多位点系统发育分析将复合种剥离既不难,也势在必行。蝉花有性型的发现是剥离这个物种复合群的第一步,今后对各大洲寄生蝉的I. cicadae近似种的研究可能会继续分解出更多的种类。
综上所述,多位点系统发育证据显示,Isaria (Paecilomyces) cicadae虽然在除南极以外的各大洲都有记载,但无论是形态还是DNA序列都表现出一定的多样性和异质性;各种证据都支持,这个当今学界广泛认可的虫草无疑是个物种复合群,其特征集要系Samson(1974)集合不同地区分布的不同近似种的特征而描述,并非同一物种在不同地区的不同标本;这样的特征集要导致了各国同行对近似种的误判。根据本研究的结果,即使在中国,在目前认定的秦岭-淮河以南的蝉花自然分布区里,被认作蝉花而使用的中药材涉及到的物种多样性也不容忽视,很可能也含有隐含种。例如,云南记载YHICYY 001和广西菌株RCEF HP090724-31的分类地位就需要进一步研究。
原产新西兰的辛克莱棒束孢Isaria sinclairii (Berk.) Lloyd被Samson(1974)组合为Paecilomyces cicadae,在世界各地都有报道,特别是日本,在东京大学综合研究博物馆就保存有大量定名为I. sinclairii的标本。遗憾的是,在GenBank中没有任何新西兰和日本的I. sinclairii序列,只能检索到Suh et al.(2016)以该名上传的ITCC 24400菌株(产地未知)的3个序列,它甚至远离虫草科而与小蝉草聚在一起,日本文化中将小蝉草与蝉花混为一谈的历史(李增智等2020)或许与此有关。东京大学的这些标本与新西兰描述的I. sinclairii是否是同一物种?与巴西物种I. cicadae是否为同一物种?本研究中韩国和葡萄牙的标本与I. cicadae是否为同一物种?再进一步,Samson(1974)组合在一起的那些亚洲种(蛁蟟棒束孢I. cosmopsaltria、蚱蝉棒束孢I. cryptotympanae)、美洲种(树棒束孢Isaria arbuscura)和非洲种(哈利奥特棒束孢Isaria harioti)与I. cicadae或I. sinclairii是否为同一物种?世界各地的I. cicadae complex可能隐含着哪些种?这些问题将在本系列论文的下一篇关于I. cicadae/I. sinclairii多样性问题的研究中进行讨论。
把虫草属Cordyceps与白僵菌属Beauveria分为两个Clade的支持率相对不高,只有69%,而虫草属内二次及三次分歧的支持率更低,分别只有48%和46%(图1)。这表明狭义虫草属(Cordyceps sensu stricto)作为单系的证据依然不足,尚需更多菌株的多位点序列证据的支持。
参考文献
Multigene phylogeny reveals new lineage for Stachybotrys chartarum, the indoor air fungus
DOI:10.1017/s0953756204000607
URL
PMID:15449591
[本文引用: 1]

Stachybotrys chartarum is an asexually reproducing fungus commonly isolated from soil and litter that is also known to occur in indoor environments and is implicated as the cause of serious illness and even death in humans. Despite its economic importance, higher level phylogenetic relationships of Stachybotrys have not been determined nor has a sexual state for S. chartarum been reported. DNA sequences from four nuclear and one mitochondrial gene were analyzed to determine the ordinal and familial placement of Stachybotrys within the Euascomycota. These data reveal that species of Stachybotrys including S. chartarum, S. albipes, for which the sexual state Melanopsamma pomiformis is reported, species of Myrothecium, and two other tropical hypocrealean species form a previously unknown monophyletic lineage within the Hypocreales. These results suggest that Stachybotrys and Myrothecium are closely related and share characteristics with other hypocrealean fungi. In addition, S. chartarum may have a sexual state in nature that consists of small, black, fleshy perithecia similar to Melanopsamma.
The Genus Cordyceps in New Zealand
Multilocus phylogenetic analyses of the genus Isaria (Ascomycota, Cordycipitaceae)
MUSCLE: multiple sequence alignment with high accuracy and high throughput
A phylogenetically-based nomenclature for Cordycipitaceae (Hypocreales)
DOI:10.5598/imafungus.2017.08.02.08
URL
PMID:29242779
[本文引用: 1]
The ending of dual nomenclatural systems for pleomorphic fungi in 2011 requires the reconciliation of competing names, ideally linked through culture based or molecular methods. The phylogenetic systematics of Hypocreales and its many genera have received extensive study in the last two decades, however resolution of competing names in Cordycipitaceae has not yet been addressed. Here we present a molecular phylogenetic investigation of Cordycipitaceae that enables identification of competing names in this family, and provides the basis upon which these names can be maintained or suppressed. The taxonomy presented here seeks to harmonize competing names by principles of priority, recognition of monophyletic groups, and the practical usage of affected taxa. In total, we propose maintaining nine generic names, Akanthomyces, Ascopolyporus, Beauveria, Cordyceps, Engyodontium, Gibellula, Hyperdermium, Parengyodontium, and Simplicillium and the rejection of eight generic names, Evlachovaea, Granulomanus, Isaria, Lecanicillium, Microhilum, Phytocordyceps, Synsterigmatocystis, and Torrubiella. Two new generic names, Hevansia and Blackwellomyces, and a new species, Beauveria blattidicola, are described. New combinations are also proposed in the genera Akanthomyces, Beauveria, Blackwellomyces, and Hevansia.
On the genus Cordyceps and its allies on Cicadidae from Japan
Tolypocladium dujiaolongae sp. nov. and its allies
Biodiversity of cordycipitoid fungi associated with Isaria cicadae Miquel, I. Nomenclatural literature study of chanhua, an important Chinese medicinal cordycipitoid mushroom recorded 1 600 years ago
A new species of Cordyceps, the telemorph of Isaria cicadae Miq
A rapid and simple extraction method for plant pathogenic fungi
On the relationships of Paecilomyces sect. Isarioidea species
New methods to calculate concordance factors for phylogenomic datasets
DOI:10.1093/molbev/msaa106
URL
PMID:32365179
[本文引用: 1]
We implement two measures for quantifying genealogical concordance in phylogenomic data sets: the gene concordance factor (gCF) and the novel site concordance factor (sCF). For every branch of a reference tree, gCF is defined as the percentage of
Sur une Novelle d’Isaria, du Brésil
Disentangling cryptic species with isaria-like morphs in Cordycipitaceae
DOI:10.1080/00275514.2018.1446651
URL
PMID:29863995
[本文引用: 1]
A new genus and eight new species, all with isaria-like phialides, are described in Cordycipitaceae from Thailand. The new genus, Samsoniella, is segregated from Akanthomyces based on morphological and molecular evidence. Samsoniella differs from Akanthomyces in producing orange cylindrical to clavate stromata with superficial perithecia and orange conidiophores with isaria-like phialides and white to cream conidia. A new combination for CBS 240.32, originally identified as Paecilomyces farinosus (Isaria farinosa), and CBS 262.58, originally identified as Penicillium alboaurantium, respectively, is made in Samsoniella. Two new species, Samsoniella aurantia and S. inthanonensis, are described from lepidopteran larvae. Two new species of Cordyceps, C. blackwelliae and C. lepidopterorum, were also found on coleopteran and lepidopteran larvae. Both produce isaria-like morphs with globose phialides and attenuated long necks and white mycelium in culture. The authors established a sexual-asexual link for Cordyceps javanica (= Isaria javanica) on lepidopteran larvae. Four new species, Akanthomyces kanyawimiae, A. sulphureus, A. thailandicus, and A. waltergamsii, were pathogenic on spiders, with some strains of A. kanyawimiae also found on unidentified insect larvae. These four species of Akanthomyces occur on the underside of leaves and produce white to cream white powdery conidia, whereas S. aurantia and S. inthanonensis were found in leaf litter and produce bright orange stromata and synnemata with white conidia. Another new combination, Akanthomyces ryukyuensis, is proposed. Phylogenetic analyses based on a combined data set comprising the nuc rDNA region encompassing the internal transcribed spacers 1 and 2 along with the 5.8S rDNA (ITS), nuc 28S rDNA (28S), partial sequences of translation elongation factor 1-alpha gene (TEF1), and the genes for RNA polymerase II largest (RPB1) and second-largest (RPB2) subunits strongly support the delimitation of these new species of Cordyceps, Akanthomyces, and in a new genus Samsoniella in Cordycipitaceae.
A Beauveria phylogeny inferred from nuclear ITS and EF1-alpha sequences: evidence for cryptic diversification and links to Cordyceps teleomorphs
DOI:10.3852/mycologia.97.1.84
URL
PMID:16389960
[本文引用: 2]
Beauveria is a globally distributed genus of soil-borne entomopathogenic hyphomycetes of interest as a model system for the study of entomopathogenesis and the biological control of pest insects. Species recognition in Beauveria is difficult due to a lack of taxonomically informative morphology. This has impeded assessment of species diversity in this genus and investigation of their natural history. A gene-genealogical approach was used to investigate molecular phylogenetic diversity of Beauveria and several presumptively related Cordyceps species. Analyses were based on nuclear ribosomal internal transcribed spacer (ITS) and elongation factor 1-alpha (EF1-alpha) sequences for 86 exemplar isolates from diverse geographic origins, habitats and insect hosts. Phylogenetic trees were inferred using maximum parsimony and Bayesian likelihood methods. Six well supported clades within Beauveria, provisionally designated A-F, were resolved in the EF1-alpha and combined gene phylogenies. Beauveria bassiana, a ubiquitous species that is characterized morphologically by globose to subglobose conidia, was determined to be non-monophyletic and consists of two unrelated lineages, clades A and C. Clade A is globally distributed and includes the Asian teleomorph Cordyceps staphylinidaecola and its probable synonym C. bassiana. All isolates contained in Clade C are anamorphic and originate from Europe and North America. Clade B includes isolates of B. brongniartii, a Eurasian species complex characterized by ellipsoidal conidia. Clade D includes B. caledonica and B. vermiconia, which produce cylindrical and comma-shaped conidia, respectively. Clade E, from Asia, includes Beauveria anamorphs and a Cordyceps teleomorph that both produce ellipsoidal conidia. Clade F, the basal branch in the Beauveria phylogeny includes the South American species B. amorpha, which produces cylindrical conidia. Lineage diversity detected within clades A, B and C suggests that prevailing morphological species concepts underestimate species diversity within these groups. Continental endemism of lineages in B. bassiana s.l. (clades A and C) indicates that isolation by distance has been an important factor in the evolutionary diversification of these clades. Permutation tests indicate that host association is essentially random in both B. bassiana s.l. clades A and C, supporting past assumptions that this species is not host specific. In contrast, isolates in clades B and D occurred primarily on coleopteran hosts, although sampling in these clades was insufficient to assess host affliation at lower taxonomic ranks. The phylogenetic placement of Cordyceps staphylinidaecola/bassiana, and C. scarabaeicola within Beauveria corroborates prior reports of these anamorph-teleomorph connections. These results establish a phylogenetic framework for further taxonomic, phylogenetic and comparative biological investigations of Beauveria and their corresponding Cordyceps teleomorphs.
Entomopathogenic fungi in Portuguese vineyards soils: suggesting a ‘Galleria-Tenebrio-bait method’ as bait-insects Galleria and Tenebrio significantly underestimate the respective recoveries of Metarhizium (robertsii) and Beauveria (bassiana)
DOI:10.3852/10-302
URL
PMID:21482632
[本文引用: 1]
Beauveria is a cosmopolitan anamorphic genus of arthropod pathogens that includes the agronomically important species, B. bassiana and B. brongniartii, which are used as mycoinsecticides for the biological control of pest insects. Recent phylogenetic evidence demonstrates that Beauveria is monophyletic within the Cordycipitaceae (Hypocreales), and both B. bassiana and B. brongniartii have been linked developmentally and phylogenetically to Cordyceps species. Despite recent interest in the genetic diversity and molecular ecology of Beauveria, particularly as it relates to their role as pathogens of insects in natural and agricultural environments, the genus has not received critical taxonomic review for several decades. A multilocus phylogeny of Beauveria based on partial sequences of RPB1, RPB2, TEF and the nuclear intergenic region, Bloc, is presented and used to assess diversity within the genus and to evaluate species concepts and their taxonomic status. B. bassiana and B. brongniartii, both which represent species complexes and which heretofore have lacked type specimens, are redescribed and types are proposed. In addition six new species are described including B. varroae and B. kipukae, which form a biphyletic, morphologically cryptic sister lineage to B. bassiana, B. pseudobassiana, which also is morphologically similar to but phylogenetically distant from B. bassiana, B. asiatica and B. australis, which are sister lineages to B. brongniartii, and B. sungii, an Asian species that is linked to an undetermined species of Cordyceps. The combination B. amorpha is validly published and an epitype is designated.
Collectotrichum gloeosporioides s.l. associated with Theobroma cacao and other plants in Panama: multilocus phylogebies distinguish host-associated pathogens from asymptomatic endophytes
DOI:10.3852/09-244
URL
PMID:20943565
[本文引用: 1]
Colletotrichum interacts with numerous plant species overtly as symptomatic pathogens and cryptically as asymptomatic endophytes. It is not known whether these contrasting ecological modes are optional strategies expressed by individual Colletotrichum species or whether a species' ecology is explicitly pathogenic or endophytic. We explored this question by inferring relationships among 77 C. gloeosporioides s.l. strains isolated from asymptomatic leaves and from anthracnose lesions on leaves and fruits of Theobroma cacao (cacao) and other plants from Panama. ITS and 5'-tef1 were used to assess diversity and to delineate operational taxonomic units for multilocus phylogenetic analysis. The ITS and 5'-tef1 screens concordantly resolved four strongly supported lineages, clades A-D: Clade A includes the ex type of C. gloeosporioides, clade B includes the ex type ITS sequence of C. boninense, and clades C and D are unidentified. The ITS yielded limited resolution and support within all clades, in particular the C. gloeosporioides clade (A), the focal lineage dealt with in this study. In contrast the 5'-tef1 screen differentiated nine distinctive haplotype subgroups within the C. gloeosporioides clade that were concordant with phylogenetic terminals resolved in a five-locus nuclear phylogeny. Among these were two phylogenetic species associated with symptomatic infections specific to either cacao or mango and five phylogenetic species isolated principally as asymptomatic infections from cacao and other plant hosts. We formally describe two new species, C. tropicale and C. ignotum, that are frequent asymptomatic associates of cacao and other Neotropical plant species, and epitypify C. theobromicola, which is associated with foliar and fruit anthracnose lesions of cacao. Asymptomatic Colletotrichum strains isolated from cacao plants grown in China included six distinct C. gloeosporioides clade taxa, only one of which is known to occur in the Neotropics.
Entomopathogenic fungi in Portuguese vineyards soils: suggesting a ‘Galleria-Tenebrio-bait method’ as baitinsects Galleria and Tenebrio significantly underestimate the respective recoveries of Metarhizium (robertsii) and Beauveria (bassiana)
Classification of Cordyceps sobolifera (Hill) Berk. et Br. and Cordyceps cicadae Shing sp. nov
Phylogenetic classification of Cordyceps and the clavicipitaceous fungi
DOI:10.3114/sim.2007.57.01
URL
PMID:18490993
[本文引用: 1]
Cordyceps, comprising over 400 species, was historically classified in the Clavicipitaceae, based on cylindrical asci, thickened ascus apices and filiform ascospores, which often disarticulate into part-spores. Cordyceps was characterized by the production of well-developed often stipitate stromata and an ecology as a pathogen of arthropods and Elaphomyces with infrageneric classifications emphasizing arrangement of perithecia, ascospore morphology and host affiliation. To refine the classification of Cordyceps and the Clavicipitaceae, the phylogenetic relationships of 162 taxa were estimated based on analyses consisting of five to seven loci, including the nuclear ribosomal small and large subunits (nrSSU and nrLSU), the elongation factor 1alpha (tef1), the largest and the second largest subunits of RNA polymerase II (rpb1 and rpb2), beta-tubulin (tub), and mitochondrial ATP6 (atp6). Our results strongly support the existence of three clavicipitaceous clades and reject the monophyly of both Cordyceps and Clavicipitaceae. Most diagnostic characters used in current classifications of Cordyceps (e.g., arrangement of perithecia, ascospore fragmentation, etc.) were not supported as being phylogenetically informative; the characters that were most consistent with the phylogeny were texture, pigmentation and morphology of stromata. Therefore, we revise the taxonomy of Cordyceps and the Clavicipitaceae to be consistent with the multi-gene phylogeny. The family Cordycipitaceae is validated based on the type of Cordyceps, C. militaris, and includes most Cordyceps species that possess brightly coloured, fleshy stromata. The new family Ophiocordycipitaceae is proposed based on Ophiocordyceps Petch, which we emend. The majority of species in this family produce darkly pigmented, tough to pliant stromata that often possess aperithecial apices. The new genus Elaphocordyceps is proposed for a subclade of the Ophiocordycipitaceae, which includes all species of Cordyceps that parasitize the fungal genus Elaphomyces and some closely related species that parasitize arthropods. The family Clavicipitaceaes. s. is emended and includes the core clade of grass symbionts (e.g., Balansia, Claviceps, Epichloe, etc.), and the entomopathogenic genus Hypocrella and relatives. In addition, the new genus Metacordyceps is proposed for Cordyceps species that are closely related to the grass symbionts in the Clavicipitaceaes. s.Metacordyceps includes teleomorphs linked to Metarhizium and other closely related anamorphs. Two new species are described, and lists of accepted names for species in Cordyceps, Elaphocordyceps, Metacordyceps and Ophiocordyceps are provided.
Sequence Matrix: concatenation software for the fast assembly of multigene datasets with character set and codon information
Ancient and recent patterns of geographic speciation in the oyster mushroom Pleurotus revealed by phylogenetic analysis of ribosomal DNA sequences
Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics
In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds.)
Resource diversity of Chinese macrofungi: edible, medicinal and poisonous species
An evaluation of common Cordyceps (Ascomycetes) species found in Chinese markets
Notes on the Medicinal Mushroom Chanhua (Cordyceps cicadae (Miq.) Massee)
多基因联合分析棒束孢属Isaria (Ascomycota,Cordycipitaceae) 系统发育关系
与蝉花有关的虫草菌生物多样性的研究I. 重要药用真菌蝉花名称的文献考证



{kind=link}
{kind=link}
{kind=link}
{kind=link}