



蜜环菌Armillaria mellea (Vahl) P. Kumm. 是伞菌目、蜜环菌属的一类真菌。在蜜环菌生活史中,菌索是常见且极其重要的组织结构,几乎所有的蜜环菌均可以形成菌索。在实验室培育或自然条件下,蜜环菌的菌索间组成一个或小或大的菌索网络,逐步向外围空间延伸,这使得蜜环菌成为地球上最大的陆地生物(Smith et al. 1992 ;Anderson & Catona 2014)。蜜环菌形成的菌索具有侵染宿主、抵御逆境、竞争其他微生物、吸收和传递营养物质、运输氧气等生物学功能(Baumgartner et al. 2011 ),这巩固了菌索作为探索性和吸收性器官的作用,适合于桥接营养源与潜在寄主之间的鸿沟。天麻和猪苓是我国传统的中药,其基源物种均可以与蜜环菌建立共生关系,蜜环菌以菌索的形式侵染二者,并为它们提供营养物质(郭顺星和徐锦堂1990,1992),蜜环菌菌索也成为影响天麻和猪苓产量的关键要素,更是评价蜜环菌菌种优劣的标准。尽管如此,蜜环菌菌丝形成菌索的相关研究较少,形成机制尚不清楚,亟待探索研究。
基于数据依赖性采集(data dependent acquisition,DDA)模式的定性蛋白质组和数据非依赖性采集(data independent acquisition,DIA)模式的SWATH定量蛋白质组学技术为挖掘差异表达蛋白提供了一种新的选择,该方法具有高效、成本相对较低、高通量、准确度高等优点,已广泛应用于动物、植物、微生物等领域,在揭示生命活动规律等方面发挥了重要作用(Collins et al. 2016 ;Li et al. 2017 )。
为探索蜜环菌菌索分化的蛋白分子机制,本研究首次对Armillaria mellea的菌丝与菌索进行了系统的蛋白质组学研究,获得了菌丝分化形成菌索的差异蛋白质组和生物信息学分析结果,为进一步研究蜜环菌提供了参考性研究方向。
1 材料与方法
1.1 材料
本研究所用菌株是本课题组从猪苓菌核中分离、培养和纯化的蜜环菌菌株,命名为A.mellea541(刘蒙蒙等 2015);菌种液氮冻存,备用。
1.2 蜜环菌培养
马铃薯琼脂糖培养基(potato dextrose agar,PDA)平皿培养A.mellea541。其固体基质菌种接种于PDA平皿中,室温避光培养。接种后的第21天收集同一皿中的蜜环菌菌丝(A.melleahypha,AH)和菌索(A.mellearhizomorphs,AR),作为对照组和实验组。样品称重,液氮速冻后超低温保存,备用。
1.3 A.mellea541菌丝形成菌索的差异蛋白质组学
A.mellea541菌丝分化形成菌索过程的差异蛋白质组方法的主要步骤包括:生物样品(菌丝和菌索)总蛋白的提取、纯化,蛋白的还原乙酰化,特异性位点的限制性酶切反应制备肽段溶液,菌丝和菌索蛋白的nano LC-Triple TOF MS/MS定性方法建立及定性数据采集,菌丝和菌索的蛋白SWATH定量方法建立及SWATH定量数据采集等,参考Huang et al.(2009) 、Li et al.(2017) 、李兵等(2017)和宋晓晶(2020)的方法,本研究也对一些参数进行了优化:优化了多肽的nano LC洗脱方法,即B相初始占比5%(A相95%)在1min内迅速升高至13%,再经69min缓慢升高B相至50%以洗脱色谱柱上的大部分肽段,后经5min过度B相至80%并等度冲洗小极性肽段5min,再经1min将B相调至初始占比5%,平衡色谱柱9min [其中:A相为甲酸-乙腈-水(0.1:2:98,V/V/V),B相为甲酸-乙腈-水(0.1:98:2,V/V/V)];高分辨模式下,母离子在线分级范围优化为300-555m/z和550-1 250m/z,Triple TOF MS的IHF设置为150,ISDF设置为2 500;SWATH定量动态采集窗口及范围包括300-323.5、322.5-352.5、351.5- 367.5m/z等60个定量采集窗口。
1.4 数据处理及生物信息学分析
1.4.1 构建A.mellea541理论蛋白库:将A.mellea541菌索和菌丝转录组的unigene序列翻译为蛋白fasta序列,以A.gallica为参比物种进行blast并用uniprot进行在线注释,获得以uniprot ID和Novel组成的A.mellea541理论蛋白库。ProteinPilot v5.0软件(AB Sciex)对以DDA模式收集的数据进行蛋白鉴定,搜索结果形成已鉴定蛋白质文库。
1.4.3 软件分析:利用OmicsbeanTM在线分析软件,对AH和AR定量数据进行质控分析,经面积归一化处理,获得A.mellea541菌所分化的差异蛋白质组(P≤0.05和AR/AH Fold change (FC) ≥1.5或≤0.067)。差异蛋白进行Gene Ontology(GO)功能注释和Kyoto Encyclopedia of Genes and Genomes(KEGG)通路分析(Ashburner et al. 2000 ;Kanehisa et al. 2016 )。
2 结果与分析
2.1 蜜环菌菌丝分化形成菌索过程的蛋白质组
A.mellea541菌丝形成菌索过程中,AH和AR总蛋白经前处理、特异性限制性酶切反应制备肽段溶液,再经除盐、富集和毛细管色谱柱洗脱后,采集的AH和AR的DDA数据经ProteinPilot分析,获得global FDR 1%范围内的二级谱图共100 185个(global FDR 1% from Fit为81.5%),匹配肽段11 627个(global FDR 1% from Fit为94.7%)。本研究从A.mellea541菌丝和菌索中共鉴定蛋白1 724个(global FDR 1%)(global FDR 1% from Fit为83.4%),其中已知蛋白1 721个(如A0A2H3DHR7、A0A2H3CTI3和A0A2H3EB94等),未知蛋白3个(包括Novel00443、Novel00043和Novel01184)。
2.2 蜜环菌菌丝形成菌索的差异蛋白质组
本研究采用非标记定量方式对A.mellea541菌丝形成菌索过程中的表达蛋白进行定量,收集菌丝和菌核样品,每组中有3个生物学重复样品,每个样品收集3次SWATH,总共有18个SWATH数据。最终有效定量蛋白1 179个,其中已知蛋白定量1 177个,未知蛋白定量2个(Novel00443和Novel00043)。
图1
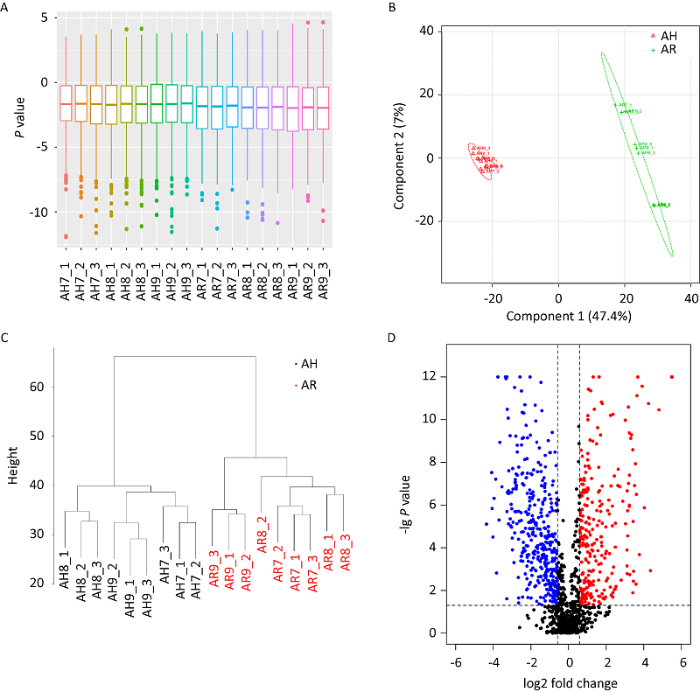
图1
蜜环菌菌丝形成菌索蛋白质组定量质控及差异蛋白分析
A:定量蛋白峰面积归一化结果;主成分分析(B)和聚类分析(C):蜜环菌菌索与菌丝聚类为两个不同的类群;D:蜜环菌菌索与菌丝间差异表达蛋白分布结果(红色为上调蛋白,蓝色为下调蛋白)
Fig. 1
Quantitative control of SWATH label-free quantitative proteome of Armillaria mellea rhizomorphs differentiated from hypha and volcano plot of differentially expressed proteins (DEPs) between rhizomorph and hypha.
A: Total quantified protein areas after normalized; B and C: Indicated rhizomorph and hypha were classed into different group by PCA and clustering analysis; D: Volcano plot of DEPs between rhizomorph group and hypha group (red blots, up-regulated proteins; blue blots, down-regulated proteins).
2.3 差异表达蛋白GO注释
A.mellea541菌丝形成菌索过程差异表达蛋白GO富集见图2。蜜环菌菌丝形成菌索中的差异表达蛋白细胞定位,即细胞组成(cell component)呈多样性分布,主要分布在细胞质和胞内细胞器。蛋白复合体是一些差异表达蛋白的广泛分布形式,包括胞内核糖体蛋白复合体、蛋白体复合体、线粒体复合体、膜蛋白复合体、ATP合酶复合体及三羧酸循环复合体等,这表明这些差异蛋白可能参与复合体组装,还参与诸如ATP合成、氧化还原、核糖体代谢等生物过程。
图2
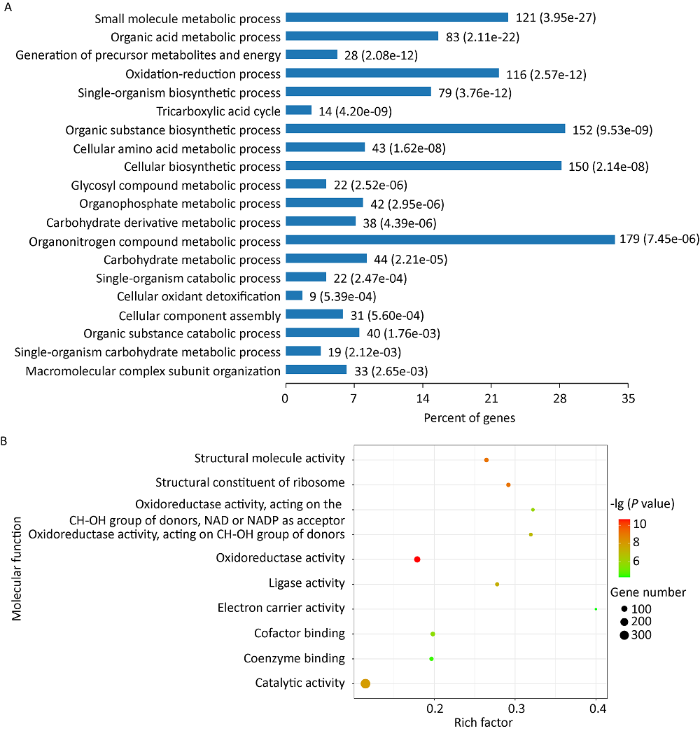
图2
蜜环菌菌丝形成菌索差异表达蛋白的GO注释富集结果
A:蜜环菌菌丝形成菌索差异表达蛋白的生物学过程分析和富集结果;B:蜜环菌菌丝形成菌索差异表达蛋白的分子功能分析和富集结果
Fig. 2
The GO analysis and enrichment results of DEPs of Armillaria mellea rhizomorphs differentiated from hypha.
A: Biological process analysis results of DEPs; B: Molecular function analysis results of DEPs.
A.mellea541菌丝形成菌索差异表达蛋白参与的生物学过程(biological process,BP)注释及富集分析见图2A。这些差异蛋白中,参与小分子代谢过程的差异蛋白占比20%,24%的差异表达蛋白与单细胞和代谢过程相关,有机含氮化合物代谢和生物合成过程的差异表达蛋白总占比17%。此外,菌丝形成菌索差异表达蛋白还参与碳水化合物代谢、蛋白定位和折叠、胞内大分子复合体聚集、内吞等生物学过程(图2A)。结果表明,菌丝分化形成菌索过程是一个复杂的生物学过程,可能涉及大分子组装、小分子代谢、蛋白表达、物质转运、氧化应激等诸多生物学途径。在差异表达蛋白中,有8个差异表达蛋白参与应答氧化应激反应,以血红素过氧化酶(heme peroxidase,A0A2H3CW41)、过氧化氢酶(catalase,A0A2H3DX77)、超氧化物歧化酶(superoxide dismutase,A0A2H3CTQ2)、OsmC样蛋白(OsmC-like protein,A0A2H3DNT3)等蛋白为代表,这些蛋白均参与氧化还原反应过程,发挥消除体内多余的活性氧物质的功能,以维持机体内活性氧物质的代谢平衡(Staerck et al. 2017 )。与菌丝相比,这些蛋白在A.mellea541菌索中上调表达(AR/AH依次为6.22、1.70、1.68和4.02),表明菌丝分化形成菌索时活性氧物质在菌索中累积,机体可能处于氧化应激状态,这反馈性地调节并激活了抗氧化反应机制。
分子功能(molecular function,MF)注释和富集分析结果也表明了蜜环菌菌丝形成菌索过程激活了氧化应激反应,以氧化还原反应富集P值最高(图2B),其中超氧化物歧化酶、过氧化氢酶、血红素过氧化酶、OsmC样蛋白等富集在抗氧化活性(antioxidant activity)通路,这再次表明了氧化应激可能在菌丝形成菌索中发挥着作用。此外,在此通路中,还有3个硫氧还蛋白样蛋白(thioredoxin-like protein,Trx),依次为A0A2H3EBV3、A0A2H3DRE2和A0A2H3E146。Trx具有清除活性氧物质发挥抗氧化作用,还是毒力蛋白,可抑制宿主免疫(Farshchi et al. 2012 ;Yang et al. 2019 )。与菌丝相比,A0A2H3E146在蜜环菌菌索中上调表达(AR/AH 2.22),而其余两种Trx下调表达(AR/AH依次为0.15和0.55),表明这3种Trx可能分属两种类型,发挥氧化或毒力因子的生物学功能。A.mellea541菌丝形成菌索过程差异表达蛋白分子功能的其余富集结果见图2B,主要包括催化活性、结构分子活性、连接酶活性、电子转运、辅酶结合活性等,其中发挥催化反应活性的差异表达蛋白数量最多,电子转运活性的富集因子最高。
2.4 差异表达蛋白KEGG通路分析
A.mellea541菌丝形成菌索过程中差异表达蛋白的KEGG通路注释及富集分析结果表明,大部分差异蛋白参与代谢过程,包括碳水化合物代谢、能量代谢、脂质代谢和氨基酸代谢等(图3)。氧化磷酸化是能量代谢的主要形式,有27个差异蛋白参与该过程。电子转移链复合体2组成蛋白琥珀酸脱氢酶铁硫亚基(succinate dehydrogenase iron-sulfur subunit,A0A2H3E088)、复合体3组成蛋白LuxS/MPP样金属水解酶(LuxS/MPP-like metallohydrolase,A0A2H3DDW6)、复合体4组成蛋白细胞色素c氧化酶亚基IV(cytochrome c oxidase subunit IV,A0A2H3E1P1)和ATP合成酶亚基e (ATP synthase subunite,A0A2H3E203)为代表的蛋白在蜜环菌菌索中上调表达,AR/AH比值依次为5.10、2.58、2.06和3.16,表明菌索ATP供能增加,能量代谢活跃(Nolfi-Donegan et al. 2020 )。参与物质代谢的差异表达蛋白数量较多,其中碳代谢富集最为显著(46个),19个蛋白与淀粉和蔗糖代谢相关、参与三羧酸循环代谢差异蛋白18个和参与糖酵解/糖异生过程的差异蛋白15个(图3的AA部分)。A.mellea541菌索形成过程中,8个差异表达蛋白参与脂肪酸代谢,7个差异表达蛋白参与酪氨酸、精氨酸、丙氨酸等的代谢(图3的BB部分)。A.mellea541菌丝形成菌索过程中,差异表达蛋白发挥的另一类功能为基因信号处理,包括核糖体代谢(37个)、氨基酰基tRNA生物合成(10个)和蛋白体代谢(15个)(图3的BC部分)。
图3
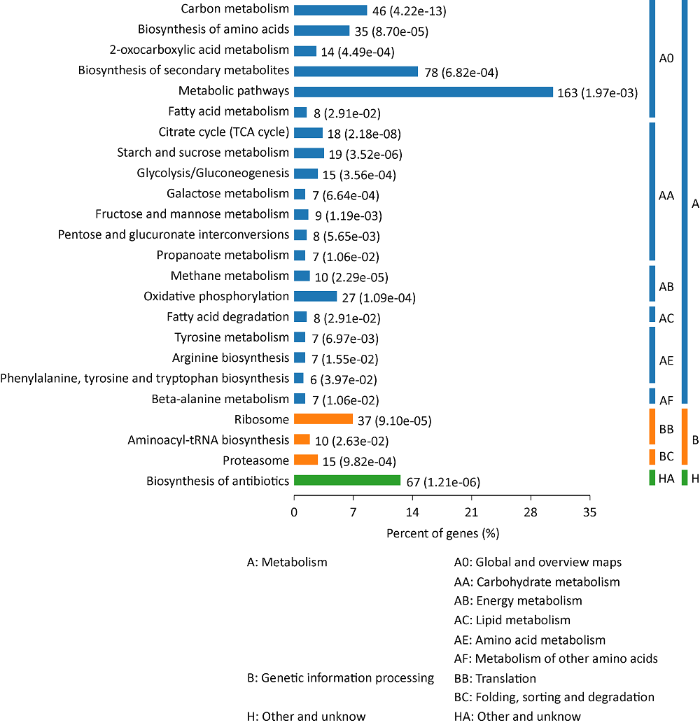
图3
蜜环菌菌丝形成菌索差异表达蛋白的KEGG注释富集结果
Fig. 3
The KEGG pathway analysis and enrichment results of DEPs of Armillaria mellea rhizomorphs differentiated from hypha.
2.5 差异表达蛋白相互作用分析
蛋白-蛋白相互作用(protein-protein interaction,PPI)分析结果见图4。在绘制的PPI网络图中,4个蛋白与碳代谢相关,为A0A2H3D6C1(富马酸还原酶,fumarate reductase)、A0A2H3E8S2(柠檬酸合酶,citrate synthase)、A0A2H3E3Z7(线粒体琥珀酸-CoA连接酶[ADP生成]β亚基,succinate-CoA ligase [ADP-forming] subunit beta)和A0A2H3EAW0(柠檬酸合酶,citrate synthase)。富马酸还原酶和线粒体琥珀酸-CoA连接酶[ADP生成] β亚基与A0A2H3E088(琥珀酸脱氢酶[泛醌]铁-硫亚基,succinate dehydrogenase [ubiquinone] iron-sulfur subunit)共同参与富马酸-琥珀酸-琥珀酰辅酶A途径的代谢,与菌丝相比,富马酸还原酶和线粒体琥珀酸-CoA连接酶[ADP生成]β亚基在菌索中下调表达(AR/AH依次为0.48和0.66)而琥珀酸脱氢酶[泛醌]铁-硫亚基上调表达(AR/AH 5.11),表明富马酸合成累积增多。此外,琥珀酸脱氢酶[泛醌]铁-硫亚基(A0A2H3E088)也是蜜环菌电子呼吸链复合体Ⅱ的亚基,在菌索中上调表达,表明菌索ATP合成增加。A0A2H3E8S2是柠檬酸代谢生成乙酰辅酶A的关键酶,在菌索中显著上调表达(AR/AH 5.59),表明代谢朝着乙酰辅酶A累积的方向进行。
图4
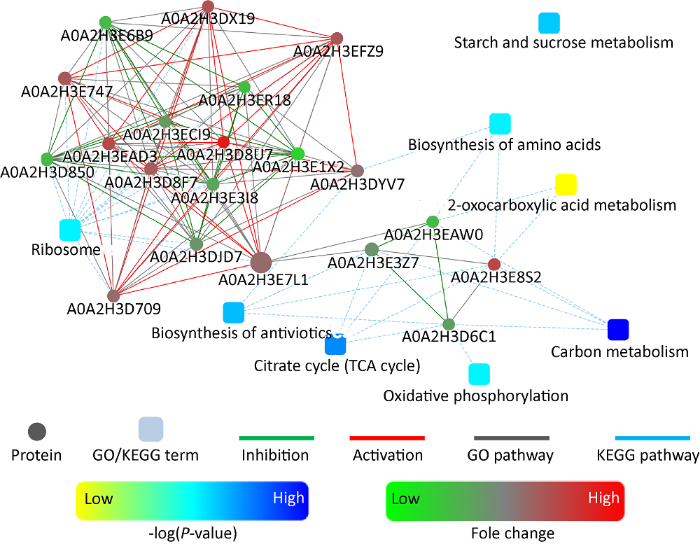
图4
蜜环菌菌丝形成菌索差异表达蛋白间相互作用分析结果
Fig. 4
The protein-protein interaction analysis and enrichment results of DEPs of Armillaria mellea rhizomorphs differentiated from hypha.
A.mellea541菌丝形成菌索过程中,碳代谢与氧化磷酸化、三羧酸循环、氨基酸生物合成、抗生素生物合成、草酰乙酸代谢和核糖体等相关联,其中以核糖体网络互作蛋白最多,共16个。泛素-40S核糖体蛋白S31融合蛋白(A0A2H3E7L1)是连接各代谢通路的枢纽蛋白,是40S核糖体的重要组成单元。敲除编码基因,则影响细胞生长(Fernández- Pevidaet al. 2016 );若该蛋白发生甲基化修饰,也可以调控盘基网柄菌细胞发育(Mangiarotti & Giorda 2002)。与A.mellea541菌丝相比,泛素-40S核糖体蛋白S31融合蛋白在菌索中上调表达(AR/AH 1.92),表明菌索的细胞生长速度可能会快于菌丝细胞的生长速度,继而菌索生长速度会快于菌丝的生长速度;无论在实验室培养还是蜜环菌伴栽猪苓或天麻的生产活动中,蜜环菌菌索的生长速度显著快于菌丝的生长速度,这表明该蛋白可能是调控蜜环菌细胞生长的一个关键蛋白。
3 讨论
蜜环菌菌索是菌丝的一种特化形式,也是侵染宿主的组织结构,菌索的多少直接影响天麻的产量(陈明义等 2004),但是蜜环菌菌丝分化形成菌索的机制目前尚不清楚。为探索蜜环菌菌索形成的机制,本研究引入先进的SWATH非标记定量蛋白质组学技术,测定了蜜环菌菌丝与菌索的蛋白质组;采用SWATH定量模式对蜜环菌菌丝和菌索中鉴定蛋白进行了非标记定量,并获得了菌丝分化形成菌索的差异表达蛋白质组。在此基础上,对差异表达蛋白质组进行了系统的GO注释、KEGG代谢通路分析和蛋白-蛋白相互作用分析,揭示了蜜环菌菌丝分化形成菌索可能是由于氧化应激所致,菌索的形成使诸如富马酸和毒力蛋白合成和释放增多,有利于蜜环菌侵染和定植于宿主。
通过对蜜环菌菌丝形成菌索差异蛋白质组的结果进行解析,表明菌索形成过程可能经历了氧化应激反应过程。活性氧是高反应性物质,如果大量累积,机体形成的氧化应激状态会对细胞成分(如核酸、脂质和蛋白质等)产生潜在的有害影响。为了将活性氧维持在无毒浓度,酶促细胞抗氧化剂可协调其生产与降解之间的平衡(Staerck et al. 2017 )。超氧化物歧化酶、过氧化氢酶、血红素过氧化酶、OsmC样蛋白和硫氧还蛋白等代表了用于清除胞内/外过量活性氧物质的重要酶类抗氧化剂。在半营养型子囊菌中,血红素过氧化物酶除具有抗氧化活性外,还可以与碳水化合物结构域功能性连接,可能对其植物致病性产生重大影响(Zámockýet al. 2017 )。超氧化物歧化酶在宿主对真菌产生的活性氧物质解毒过程中发挥着重要的作用。超氧化物歧化酶和过氧化氢酶可清除真菌体内的超氧负离子和过氧化氢,或者抵御宿主产生的活性氧物质,以维持机体活性氧物质平衡和发挥免疫原活性(Shibuya et al. 2006 ;Chaves & da Silva 2012)。对于一些致病性真菌,超氧化物歧化酶也被认为是一种细胞毒力因子(Hwang et al. 2002 ),如白色念珠菌。在大肠杆菌中,OsmC样蛋白利用其高反应性半胱氨酸硫醇基团清除过氧化产物(Lesniak et al. 2003 )。硫氧还蛋白是抗氧化酶系中的另一类蛋白,硫氧还蛋白样蛋白是与硫氧还蛋白具有高度保守结构域的蛋白,敲除肺炎球菌中编码该蛋白的基因(Etrx1),肺炎球菌对活性氧物质的敏感性降低(Farshchi Andisi et al. 2012 )。A.mellea541菌丝形成菌索过程有3个硫氧还蛋白样蛋白差异表达,其中硫氧还蛋白样蛋白(A0A2H3E146)在菌索中上调表达,说明菌索对活性氧物质的敏感性增加,抗氧化能力增强。另据Yang et al.(2019) 报道,爱德华氏菌Edwardsiella piscicida是一种鱼类寄生菌,可以释放硫氧还蛋白样蛋白,该蛋白可作为细胞毒力因子,消除宿主的ASK1(apoptosissignal-regulating kinase 1)的同源相互作用和磷酸化,抑制下游胞外信号调节蛋白激酶——p38-丝裂原活化蛋白激酶(extracellular signal-regulated kinase 1/2-p38-mitogen-activated protein kinase,Erk1/2-p38-MAPK) 信号通路而抵御宿主免疫反应,以达到侵染的目的。猪苓或天麻的生长发育离不开蜜环菌,蜜环菌以菌索的组织形式侵染二者,最终建立共生关系。这些蛋白在蜜环菌菌索中上调表达,一方面说明氧化应激可能诱导菌丝分化继而形成菌索,另一方面菌索释放毒力因子并可能通过Erk1/2-p38-MAPK信号通路途径抑制宿主免疫反应而实现侵染的目的,其具体机制有待深入研究。
在蜜环菌菌丝分化形成菌索的过程中,与菌丝相比,电子呼吸链复合体(含ATP合成酶)的一些亚基和富马酸代谢酶在蜜环菌菌索中上调表达,这可能产生两种结果,一方面ATP合成增加,供能充足(Fernie et al. 2004 ),更有利于菌索的形成、分枝甚至去侵染宿主;另一方面,累积的有机酸(富马酸)或许可能释放到环境,溶解矿物质,增加菌索吸收和转运如磷等类物质的能力;此外,过多的释放富马酸到环境中,也改变了宿主的生活条件,累积的富马酸也可以抑制宿主的线粒体代谢(Schliemann et al. 2008 ),从而增加蜜环菌菌索侵染宿主的几率。蜜环菌是宿主(如天麻、猪苓菌核)的营养供给体,为它们的生长发育提供充足的营养物质(郭顺星和徐锦堂1990,1992),有机酸的释放或许加快蜜环菌与宿主共生关系的建立,而蜜环菌菌索的形成促进了这一进程。
参考文献
Genomewide mutation dynamic within a long-lived individual of Armillaria gallica
DOI:10.3852/13-367
PMID:24891414
[本文引用: 1]

Mutation is the ultimate source of all genetic variation in populations and yet they remain unobservable and buried in the past. Long-lived individuals of Armillaria gallica, a common opportunistic fungal pathogen of tree roots in temperate forests of the northern hemisphere, provide a spatial context for examining the mutational dynamic. Each individual of A. gallica arises in a single mating between two haploid gametes and the resulting diploid then grows vegetatively to occupy a discrete spatial territory often including many adjacent tree root systems. In effect, this leaves a spatial record of growth over time within which mutations can be localized. To identify mutations, the entire genomes of three spatially separated samples of one individual of A. gallica approximately 200 × 60 m were sequenced and compared. In this comparison, mutations and regions of loss of heterozygosity (LOH) were identified then assayed in another 20 isolates from the same individual by conventional PCR and Sanger sequencing. The genotype network of all mutations and LOH were without internal conflict. Further, the spatial pattern of genotypes was nonrandom and appeared to reflect the vegetative expansion leading to the present-day individual. The results reflect the spectrum of spontaneous mutation in nature and provide insight into cellular generation times. © 2014 by The Mycological Society of America.
Gene ontology: tool for the unification of biology
DOI:10.1038/75556 URL [本文引用: 1]
Secrets of the subterranean pathosystem of Armillaria
DOI:10.1111/mpp.2011.12.issue-6 URL [本文引用: 1]
Superoxide dismutases and glutaredoxins have a distinct role in the response of Candida albicans to oxidative stress generated by the chemical compounds menadione and diamide
DOI:10.1590/S0074-02762012000800006 URL [本文引用: 1]
Effect of different strains of Armillariella mellea on the yield of Gastrodia elata
Multi-laboratory assessment of reproducibility, qualitative and quantitative performance of SWATH-mass spectrometry
DOI:10.1038/s41467-017-00249-5 URL [本文引用: 1]
Pneumococcal gene complex involved in resistance to extracellular oxidative stress
DOI:10.1128/IAI.05563-11 URL [本文引用: 2]
The eukaryote-specific N-terminal extension of ribosomal protein S31 contributes to the assembly and function of 40S ribosomal subunits
DOI:10.1093/nar/gkw641 URL [本文引用: 1]
Respiratory metabolism: glycolysis, the TCA cycle and mitochondrial electron transport
Studies on the cell ultrastructrue in the course of Gastrodia elata digesting Mycena osmundicola Lange and Armillaria mellea Fr
Nutrient source of sclerotia of Grifola umbelata and its relationship to Armillaria mellea
Direct identification and quantification of aspartyl succinimide in an IgG2 mAb by RapiGest SF assisted digestion
DOI:10.1021/ac802708s URL [本文引用: 1]
Copper-andzinc-containing superoxide dismutase (Cu/ZnSOD) is required for the protection of Candida albicans against oxidative stresses and the expression of its full virulence
DOI:10.1099/00221287-148-11-3705 URL [本文引用: 1]
KEGG as a reference resource for gene and protein annotation
Structural and functional features of the Escherichia coli hydroperoxide resistance protein OsmC
The osmotically inducible protein OsmC, like its better-characterized homolog, the organic hydroperoxide protein Ohr, is involved in defense against oxidative stress caused by exposure to organic hydroperoxides. The crystal structure of Escherichia coli OsmC reported here reveals that the protein is a tightly folded domain-swapped dimer with two active sites located at the monomer interface on opposite sides of the molecule. We demonstrate that OsmC preferentially metabolizes organic hydroperoxides over inorganic hydrogen peroxide. On the basis of structural and enzymatic similarities, we propose that the OsmC catalytic mechanism is analogous to that of the Ohr proteins and of the structurally unrelated peroxiredoxins, directly using highly reactive cysteine thiol groups to elicit hydroperoxide reduction.
SWATH label-free proteomics analyses revealed the roles of oxidative stress and antioxidant defensing system in sclerotia formation of Polyporus umbellatus
DOI:10.1038/srep41283 URL [本文引用: 2]
Differential proteome analysis of sclerotium formation stages of Polyporus umbellatus
Investigation of symbioticArmillaria species with Chinese traditional medicinal fungus Polyporus umbellatus
Synthesis of ribosomal proteins in developing Dictyostelium discoideum cells is controlled by the methylation of proteins S24 and S31
Ribosomal protein mRNAs left over from growth are selectively excluded from polyribosomes in the first half of Dictyostelium discoideum development. This is due to the fact that they are sequestered by a class of free 40S ribosomal subunits, characterized by possessing a methylated S24 protein. At the time of formation of tight cell aggregates, the methylated S24 is substituted by an unmethylated S24, while protein S31 of the same or other 40S subunits becomes methylated. This leads to a rapid degradation of the ribosomal protein mRNAs.
Mitochondrial electron transport chain: oxidative phosphorylation, oxidant production, and methods of measurement
DOI:10.1016/j.redox.2020.101674 URL [本文引用: 1]
Metabolite profiling of mycorrhizal roots of Medicago truncatula
Metabolite profiling of soluble primary and secondary metabolites, as well as cell wall-bound phenolic compounds from roots of barrel medic (Medicago truncatula) was carried out by GC-MS, HPLC and LC-MS. These analyses revealed a number of metabolic characteristics over 56 days of symbiotic interaction with the arbuscular mycorrhizal (AM) fungus Glomus intraradices, when compared to the controls, i.e. nonmycorrhizal roots supplied with low and high amounts of phosphate. During the most active stages of overall root mycorrhization, elevated levels of certain amino acids (Glu, Asp, Asn) were observed accompanied by increases in amounts of some fatty acids (palmitic and oleic acids), indicating a mycorrhiza-specific activation of plastidial metabolism. In addition, some accumulating fungus-specific fatty acids (palmitvaccenic and vaccenic acids) were assigned that may be used as markers of fungal root colonization. Stimulation of the biosynthesis of some constitutive isoflavonoids (daidzein, ononin and malonylononin) occurred, however, only at late stages of root mycorrhization. Increase of the levels of saponins correlated AM-independently with plant growth. Only in AM roots was the accumulation of apocarotenoids (cyclohexenone and mycorradicin derivatives) observed. The structures of the unknown cyclohexenone derivatives were identified by spectroscopic methods as glucosides of blumenol C and 13-hydroxyblumenol C and their corresponding malonyl conjugates. During mycorrhization, the levels of typical cell wall-bound phenolics (e.g. 4-hydroxybenzaldehyde, vanillin, ferulic acid) did not change; however, high amounts of cell wall-bound tyrosol were exclusively detected in AM roots. Principal component analyses of nonpolar primary and secondary metabolites clearly separated AM roots from those of the controls, which was confirmed by an hierarchical cluster analysis. Circular networks of primary nonpolar metabolites showed stronger and more frequent correlations between metabolites in the mycorrhizal roots. The same trend, but to a lesser extent, was observed in nonmycorrhizal roots supplied with high amounts of phosphate. These results indicate a tighter control of primary metabolism in AM roots compared to control plants. Network correlation analyses revealed distinct clusters of amino acids and sugars/aliphatic acids with strong metabolic correlations among one another in all plants analyzed; however, mycorrhizal symbiosis reduced the cluster separation and enlarged the sugar cluster size. The amino acid clusters represent groups of metabolites with strong correlations among one another (cliques) that are differently composed in mycorrhizal and nonmycorrhizal roots. In conclusion, the present work shows for the first time that there are clear differences in development- and symbiosis-dependent primary and secondary metabolism of M. truncatula roots.
Catalases of Aspergillus fumigatus and inflammation in aspergillosis
DOI:10.3314/jjmm.47.249 URL [本文引用: 1]
The fungus Armillaria bulbosais among the largest and oldest living organisms
DOI:10.1038/356428a0 URL [本文引用: 1]
The studies on low hydraulic resistanc channels along meridians in rats and its morphology and differential proteome
Microbial antioxidant defense enzymes
DOI:10.1016/j.micpath.2017.06.015 URL [本文引用: 2]
The Edwardsiella piscicida thioredoxin- like protein inhibits ASK1-MAPKs signaling cascades to promote pathogenesis during infection
DOI:10.1371/journal.ppat.1007917 URL [本文引用: 2]
Fungal hybrid B heme peroxidases-unique fusions of a heme peroxidase domain with a carbohydrate- binding domain
DOI:10.1038/s41598-017-09581-8 URL [本文引用: 1]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}