



微小染色体维持蛋白(minichromosome maintenance protein,Mcm)是DNA复制启动和延伸的关键因素(Juríková et al. 2016),也是保证一个细胞周期(cell cycle)内仅发生一次染色体复制的重要许可因子(Tye 1999;Bochman & Schwacha 2009;Juríková et al. 2016;Zhai et al. 2017)。Mcm家族中Mcm2、Mcm3、Mcm4、Mcm5、Mcm6、Mcm7组成的六聚体复合物(Mcm2-7)是真核生物DNA复制解旋酶的核心(Matson et al. 2017;Zhai et al. 2017),它们能将亲代DNA双螺旋链解旋,辅助聚合酶合成新的DNA(Bleichert et al. 2017),进而保证了生物细胞周期的顺利进行。
目前,Mcm报道主要集中在癌症方面,研究发现在不同肿瘤中呈现异常表达并与肿瘤细胞增殖、侵袭、转移能力密切相关。Mcm6是神经胶质瘤病人预后不良的潜在标志物(Cai et al. 2018),是食管鳞状细胞癌治疗和诊断的标志物,Mcm6基因敲除抑制了细胞增殖、迁移和侵染,促进了细胞凋亡,并将细胞阻滞在S期(Li et al. 2020)。Mcm2基因沉默能显著阻滞肝癌细胞HepG2细胞周期,并抑制其增殖率(Yang et al. 2018)。敲除Mcm4基因表达可以减少白血病细胞的比例(Peng et al. 2018)。这些研究表明Mcm蛋白可以作为临床上诊断相关恶性肿瘤及提示预后的生物学标志物,更为重要的是Mcm作为靶基因可以为新型抗肿瘤药物的研发提供新思路。
葡萄座腔菌Botryosphaeria dothidea引起的苹果轮纹病是我国苹果产业中的三大病害之一(侯珲等 2017)。据调查在我国7个主产区,苹果枝干轮纹病的总体发病率高达77.6%(国立耘等 2009)。该病危害果树枝干,引起“粗皮病”,造成果树树势衰弱,影响果树产量及果品品质。而且,葡萄座腔菌侵染果实,在成熟期或者贮藏期引起果实腐烂,难以控制。每年因该病导致的果实腐烂约10%-20%,最高可达70%(Zhao et al. 2016)。我国是世界上最大的苹果生产国,面积、产量均占世界的50%以上,年总产值达2 100亿元。“富士”是我国苹果主栽品种,约占苹果产业的70%(陈学森等 2020)。而“富士”易感轮纹病,发病尤为严重(侯珲等 2017)。因此,苹果轮纹病对我国苹果产业造成巨大影响,严重危害苹果产业的健康持续发展,亟待探明其致病机理并研发高效安全的防控措施。
前期研究也表明,韭菜提取液及其中所含的有机硫化物能显著抑制葡萄座腔菌的生长(赵刚等2017)。那么葡萄座腔菌Mcm基因家族能是否能响应硫化物胁迫,能否作为解析苹果轮纹病病菌的发病机理的重要因素?能否作为研发防控苹果轮纹病的潜在靶标基因?为了明确这个问题,本研究在全基因组水平鉴定了葡萄座腔菌Mcm基因家族,并分析了其在硫化物胁迫下的表达特性,以期为通过分子手段防控该病提供理论基础。
1 材料与方法
1.1 材料的制备
葡萄座腔菌培养于PDA培养基上,保存在本实验室。硫化物二甲基三硫购自成都麦卡西公司。取20μL液体PDA培养基倒入直径9cm的培养皿,冷却后,铺上玻璃纸,在玻璃纸中央接种一枚直径为0.5cm的葡萄座腔菌饼,倒置于28℃培养箱中培养。2d后,在培养皿上盖中央放置一小块无菌滤纸,取125μL/L的二甲基三硫100μL滴加到滤纸上,封口,倒置在28℃培养箱中继续培养1h后,刮取菌丝备用。
1.2 葡萄座腔菌Mcm基因家族的获得
在NCBI下载葡萄座腔菌基因组数据(ASM1150312v2)(Yu et al. 2020),然后在Ensemble(
1.3 葡萄座腔菌Mcm基因家族基本特性分析
利用在线软件expasy(
1.4 葡萄座腔菌Mcm基因家族表达特性
本实验室前期发布了硫化物二甲基三硫醚处理1h后的葡萄座腔菌RNA-seq数据(保存于国家微生物科学数据中心NMDC:
表1 葡萄座腔菌Mcm基因家族qRT-PCR引物
Table 1
| 基因 Gene | 上游引物 Forward primer sequence (5′-3′) | 下游引物 Reverse primer sequence (5′-3′) |
|---|---|---|
| BdMcm2 | GTAAAGTCGCTCAGGGCAAG | AGCGTCCACTCGCTAGTCAT |
| BdMcm3 | GAAGACGAGAACGGAAGTGC | GTCAAATTTGCGTGGTTGTG |
| BdMcm4 | GCAGCAAGTACAACCCCAAT | GAGGTACATGCCGACCAGAT |
| BdMcm5 | CGCAACGAGAATGAGGACAG | TGTGCCAGCTCCTCATTGTA |
| BdMcm6 | CTAACCAGACTGGCGAGGAG | TTGATGACCTGGTGTCCGTA |
| BdMcm7 | AAAGCTGGAATCACCACCAC | GGTTTCAGGGTGCTTGTTGT |
| Actin | GTTCAGACCGCCCTTTGCT | AGCCTTGCGACGGAACATA |
1.5 细胞周期检测
取1.1部分准备的鲜菌丝1g于50mL的离心管中,加入10mL 0.6%酶解液(溶壁酶:崩溃酶=1:1),在28℃、100r/min的摇床上振荡酶解3h,用0.7mol/L的NaCl溶液冲洗2次,4℃,3 000r/min离心10min,弃上清液,沉淀即为原生质体。用1mL的STC溶液悬浮原生质体,并加于2mL无菌离心管中,按照真菌/酵母细胞样品细胞周期碘化丙啶红色荧光流式细胞分析试剂盒(GenMed Scientifics Inc. US)方法处理样品。首先加入1mL清理液(Reagent A),1 200r/min离心1min,去上清液;缓慢加入2mL预冷的固着液(Reagent B),4℃冰箱里孵育16h,1 200r/min离心1min,去上清液;重复上述步骤后,加入500μL清液抗干扰液(Reagent C),37℃恒温孵育16h,1 200r/min离心1min,去上清液;加入200μL清液酶解液(Reagent D)37℃恒温孵育20min,1 200r/min离心1min,去上清液;加入500μL清液清理液(Reagent A),1 200r/min离心1min,去上清液后,再加入500μL清液染色液(Reagent E),超声处理15s,暗室37℃孵育30min,4℃冰箱里待测。细胞周期利用流式细胞仪(FACS Calibur,美国BD公司)进行检测,流式细胞仪光源为488nm氩离子激光,FL3荧光通道进行检测,收集10 000个细胞。每个处理3次重复,利用FlowJo v10软件(FlowJo LLC.,USA)分析细胞周期各阶段细胞百分数。
2 结果与分析
2.1 葡萄座腔菌Mcm基因家族基本特性分析
通过在葡萄座腔菌基因组中搜索Mcm基因家族,得到BOTSDO10608.1、BOTSDO10478.1、BOTSDO10560.1、BOTSDO01390.1、BOTSDO 03037.1、BOTSDO09452.1和BOTSDO07461.1等7条基因序列。在NCBI CDD进行保守结构域分析显示,BOTSDO07461.1并不具备Mcm-box保守结构域,而具有MADS-box保守结构域。通过hmmer分析从葡萄座腔菌中共鉴定到6条基因序列,BOTSDO10608.1、BOTSDO10478.1、BOTSDO10560.1、BOTSDO01390.1、BOTSDO 09452.1、BOTSDO03037.1。将所有候选序列合并去重,得到6条序列。在NCBI CDD进行保守结构域分析显示,6条基因均含有Mcm-box保守结构域。因此,认定这6条基因即是葡萄座腔菌Mcm基因(BdMcm),将其命名为BdMcm2-BdMcm7。BdMcm2- BdMcm7位于5个重叠群(Contig)上,其中BdMcm3、BdMcm5、BdMcm6和BdMcm7分别位于重叠群73、重叠群1、重叠群62、重叠群16上,而BdMcm2和BdMcm4共同位于重叠群82上。BdMcm2-BdMcm7编码产物长度为718(BdMcm5)-1 618(BdMcm3)个氨基酸,平均长度为994.17个氨基酸。其等电点为5.04(BdMcm6)-8.33(BdMcm4),平均为6.13。而分子量在79.78kDa(BdMcm5)-180.30kDa(BdMcm3),平均为110.25kDa(表2)。
表2 葡萄座腔菌Mcm基因家族基本理化特性
Table 2
| 基因 Gene | 基因ID号 Gene ID | 重叠群 Contig | 染色体上位置 Location on the chromosome | 蛋白长度 Length (aa) | 蛋白等电点PI | 蛋白分子量 MW (kDa) |
|---|---|---|---|---|---|---|
| BdMcm2 | BOTSDO10608.1 | WWBZ02000082.1 | 283 862-286 547 | 852 | 5.13 | 95.37 |
| BdMcm3 | BOTSDO10478.1 | WWBZ02000073.1 | 3 359 906-3 365 858 | 1 618 | 6.55 | 180.30 |
| BdMcm4 | BOTSDO10560.1 | WWBZ02000082.1 | 125 412-128 550 | 1 010 | 8.33 | 110.53 |
| BdMcm5 | BOTSDO01390.1 | WWBZ02000001.1 | 4 682 589-4 684 847 | 718 | 6.40 | 79.78 |
| BdMcm6 | BOTSDO09452.1 | WWBZ02000062.1 | 3 397 693-3 400 684 | 958 | 5.04 | 105.30 |
| BdMcm7 | BOTSDO03037.1 | WWBZ02000016.1 | 4 197 732-4 200 376 | 809 | 5.38 | 90.21 |
2.2 葡萄座腔菌Mcm基因家族进化分析及分类
为了对葡萄座腔菌Mcm基因家族的6个成员进行分类,将其与拟南芥、构巢曲霉、人、水稻、酵母、苹果腐烂病菌共7个物种的Mcm基因家族共同进行聚类。结果显示,7个物种的Mcm基因家族分为8个亚类,分别对应为Mcm2、Mcm3、Mcm4、Mcm5、Mcm6、Mcm7、Mcm8、Mcm9(图1)。其中,BdMcm2-BdMcm7分别位于相应的亚类中。而且,在每个亚类中,葡萄座腔菌Mcm基因家族与同为真菌的构巢曲霉Mcm、苹果腐烂病菌Mcm、酵母Mcm基因家族关系较近。而水稻、拟南芥和人除具有Mcm2-Mcm7外,还有具有Mcm8和Mcm9。利用clustal X将BdMcm2-BdMcm7比对发现,它们均含有Mcm结构域的特征序列,Walker A(GDPXX(S/A)KS)、Walker B(IDEFDKM),以及R-finger(SRFD)(图2)。
图1
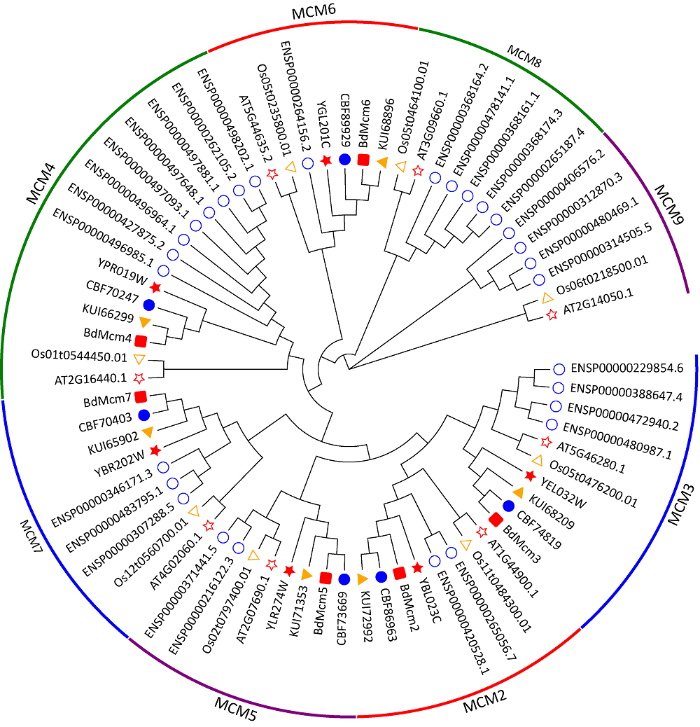
图1
葡萄座腔菌等7个物种的Mcm基因家族的进化树分析







Fig. 1
Phylogenetic tree constructed by the Mcm gene families from seven species.


图2

图2
葡萄座腔菌Mcm基因家族保守结构域序列比对
Fig. 2
Sequence alignment of conserved domains in Mcm gene family of Botryosphaeria dothidea.
2.3 葡萄座腔菌Mcm基因家族结构域、motif及基因结构分析
在NCBI CDD进行葡萄座腔菌Mcm基因家族保守结构域分析可以看出,BdMcm2- BdMcm7都含有Mcm_N和Mcm结构域,另外BdMcm2含有一个Mcm2_N,BdMcm6含有一个特殊的Mcm6_C(图3A)。对葡萄座腔菌Mcm基因家族motif分析发现,BdMcm2- BdMcm7都含有motif1、motif2、motif3、motif4、motif5、motif6、motif7和motif8(表3),BdMcm2、BdMcm3、BdMcm4、BdMcm5含有特殊的motif9,而BdMcm4和BdMcm6含有特殊的motif10。其中motif1、motif2、motif3、motif4和motif8为Mcm保守结构域(图3B)。葡萄座腔菌Mcm基因家族基因结构分析显示BdMcm2、BdMcm4、BdMcm5、BdMcm6含有3个CDS,BdMcm7含有5个CDS,而BdMcm3最多,有14个CDS(图3C)。
图3
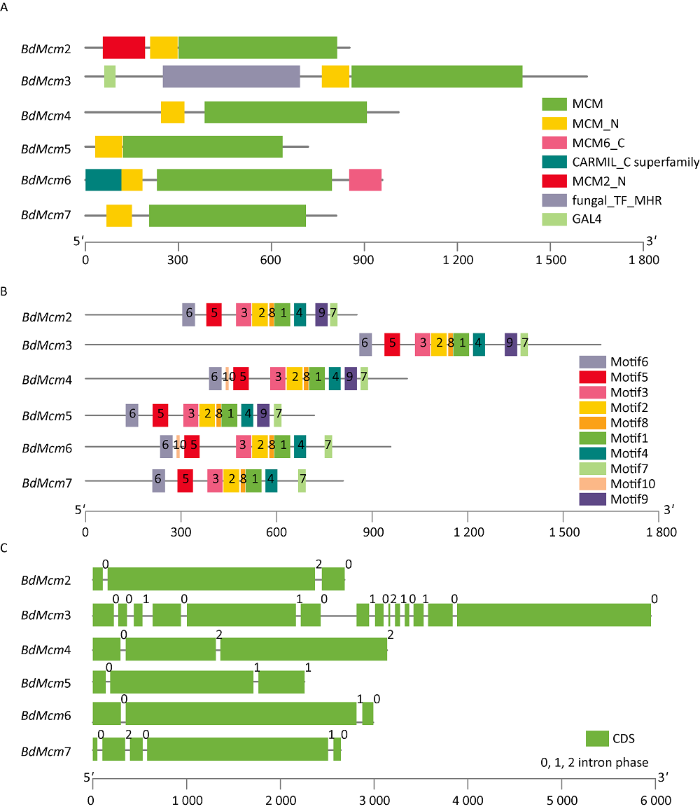
图3
葡萄座腔菌Mcm基因家族保守结构域(A)、基序(B)和基因结构(C)
Fig. 3
Domains (A), motifs (B) and gene structures (C) of Mcm gene family of Botryosphaeria dothidea.
表3 葡萄座腔菌Mcm基因家族motif序列
Table 3
| Motif | Width | Sequence |
|---|---|---|
| 1 | 50 | CCIDEFDKMDDADRVAIHEVMEQQTISIAKAGITTTLNARTSILAAANPI |
| 2 | 50 | RGDINVLLLGDPGTAKSQLLKYVEKVAPRAVYTSGKGSSAVGLTAAVTRD |
| 3 | 48 | IRELSKRPDIYELLSRSIAPSIYGHDDIKKAILLQLFGGVSKELPDGM |
| 4 | 38 | NIBLPAPJLSRFDLLFVVLDEIDEGRDERLARHVVGIH |
| 5 | 49 | FRDYQKIKJQETPDZVPVGQLPRHVDVILDGDLVDRVKPGDRVEITGIF |
| 6 | 40 | RQLRSSHLNHLVSISGIVTRTSEVRPKLKAAAFSCENCNV |
| 7 | 24 | YPITVRQLESIIRJSEAHAKMRLS |
| 8 | 15 | EFVLEGGALVLADGG |
| 9 | 39 | EJPVEFIKKYISYAKSKVKPRLTQAAAEKJVSEYVGMRR |
| 10 | 10 | EPTQCPRPIC |
2.4 葡萄座腔菌Mcm基因家族亚细胞定位预测
将BdMcm2-BdMcm7蛋白序列利用ProtComp Version 9.0软件对其亚细胞定位进行预测。结果显示,葡萄座腔菌Mcm基因家族6个成员均主要定位在细胞核内。其中BdMcm2、BdMcm3、BdMcm4、BdMcm6在高尔基体内有少量分布,BdMcm4、BdMcm5、BdMcm6和BdMcm7在细胞质膜中也有一定量的表达。
2.5 葡萄座腔菌Mcm基因家族表达分析
从硫化物二甲基三硫醚处理葡萄座腔菌1h后的RNA-seq数据中提取葡萄座腔菌Mcm基因家族的表达量(FPKM)分析发现,与对照相比,葡萄座腔菌Mcm基因家族6个成员的表达量均明显下降,BdMcm2、BdMcm3、BdMcm4、BdMcm5、BdMcm6、BdMcm7分别比对照降低75.53%、20.56%、72.14%、78.10%、61.60%、75.47%(图4)。
图4
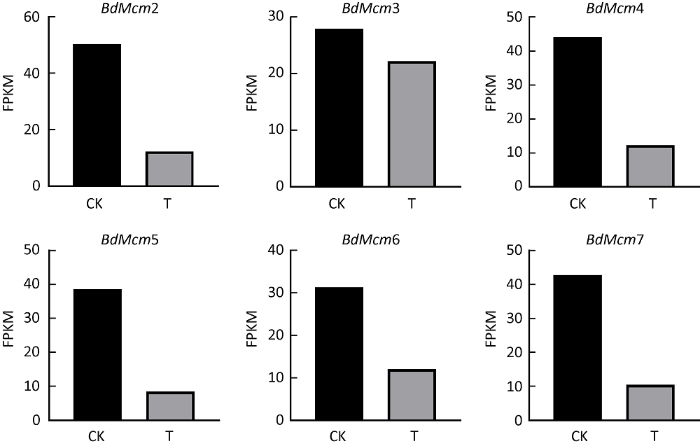
图4
二甲基三硫醚处理1h后,通过RNA-seq方法检测葡萄座腔菌Mcm基因家族表达量
CK:对照;T:处理
Fig. 4
The relative expression (FPKM) of Mcm gene family in Botryosphaeria dothidea detected by RNA-seq method after 1h treatment with dimethyl trithioether.
CK: Control; T: Treatment.
为了进一步验证硫化物葡萄座腔菌Mcm基因表达的影响,通过qRT-PCR方法检测了硫化物二甲基三硫醚胁迫处理葡萄座腔菌1h后葡萄座腔菌Mcm基因家族6个成员表达情况。结果显示,与对照相比,6个BdMcm基因表达水平呈现不同程度下降趋势。BdMcm4表达量虽下降10.74%,但与对照相比,并不显著(F=4.3,P=0.1068)。除此之外,其他5个BdMcm基因都呈显著或者极显著下降趋势,其中BdMcm2、BdMcm3、BdMcm5、BdMcm6、BdMcm7表达量分别比对照下降21.27%、73.83%、50.56%、17.65%、67.90%(图5)。相关性分析表明,RNA-seq数据中提取的葡萄座腔菌Mcm基因表达量与qRT-PCR检测的相对表达量变化趋势一致,二者具有高度相关性(R2=0.8958)(图6)。
图5
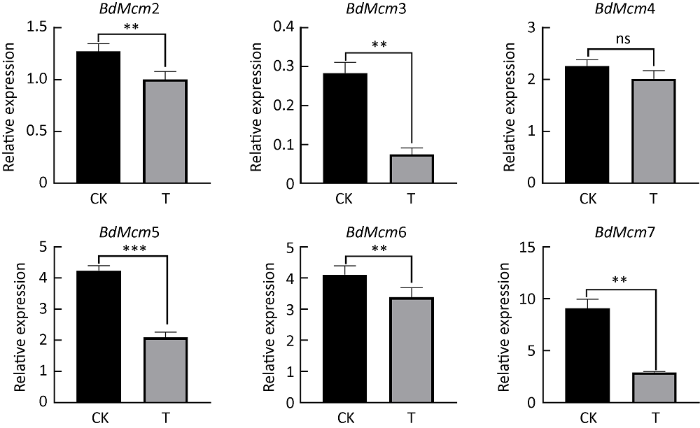
图5
二甲基三硫醚处理1h后,通过荧光定量PCR方法检测葡萄座腔菌Mcm基因家族表达量
CK:对照;T:处理. ns表示P>0.05,*表示0.05<P<0.01,**表示0.0001<P<0.01,***表示P<0.0001
Fig. 5
The relative expression of Mcm gene family in Botryosphaeria dothidea detected by quantitative real-time PCR method after 1h treatment with dimethyl trithioether.
CK: Control; T: Treatment. ns represented P>0.05, * represented 0.05<P<0.01, ** represented 0.0001<P<0.01, and *** represented P<0.0001.
图6
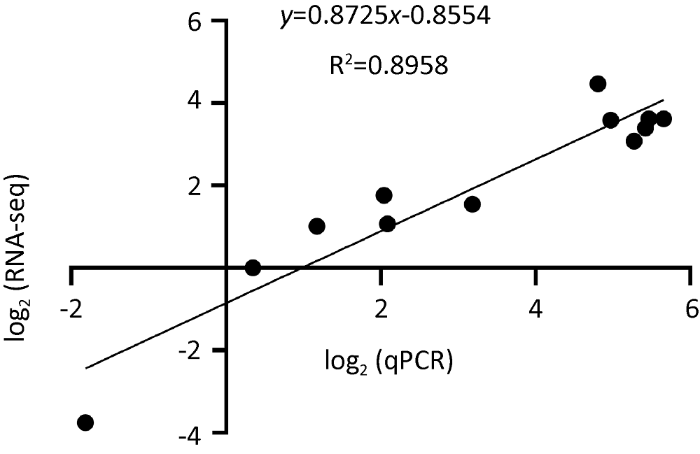
图6
RNA-seq和荧光定量PCR检测葡萄座腔菌中Mcm基因家族表达量的相关性分析
Fig. 6
Correlation analysis of Mcm gene family expression in Botryosphaeria dothidea detected by RNA-seq and quantitative real-time PCR.
2.6 硫化物阻滞了葡萄座腔菌细胞周期
通过流式细胞仪检测了硫化物二甲基三硫醚处理1h后葡萄座腔菌细胞周期的情况。结果表明,硫化物显著阻滞了其正常的细胞周期(图7)。对照菌中G0/G1期细胞为31.2%,S期的细胞为48.1%,G2/M期的细胞为20.7%。与之相比,硫化物处理后,G0/G1期细胞提高了45.3%,(F=9.14,P=0.039),G2/M期细胞比例提高了17.5%,但并不显著(F=1.46,P=0.2931),而S期细胞比例下降了37.0%(F=70.51,P=0.0011),说明硫化物处理在G0/G1期显著阻滞了细胞周期进程。
图7
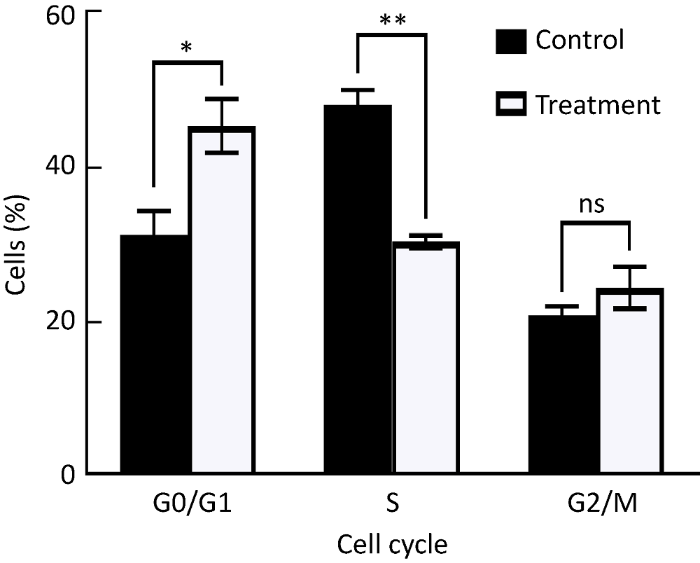
图7
二甲基三硫醚对葡萄座腔菌细胞周期的影响
ns表示P>0.05,*表示0.05<P<0.01,**表示0.0001<P<0.01
Fig. 7
The effects of dimethyl trithioether on the cell cycle of Botryosphaeria dothidea.
ns represented P>0.05, * represented 0.05<P<0.01, and ** represented 0.0001<P<0.01.
3 讨论
目前已经鉴定到10个Mcm基因家族成员,即Mcm1-Mcm10(王东星等 2018)。研究表明,Mcm1与Mcm10功能上与Mcm蛋白相似,但是不属于此类蛋白家族(Forsburg 2004)。而Mcm8和Mcm9仅存在于高等生物体内(付凌玲等 2018)。实际上真正的Mcm基因家族只有Mcm2-Mcm7(Forsburg 2004)。本研究从葡萄座腔菌基因组中提取的BOTSDO07461.1序列,经分析得知其具有MADS-box保守结构域,而并不具有Mcm-box保守结构域,实为BdMcm1基因家族。随后利用hmmer方法在葡萄座腔菌基因组中重新鉴定,也只鉴定得到6个葡萄座腔菌Mcm基因家族成员,即BdMcm2、BdMcm3、BdMcm4、BdMcm5、BdMcm6和BdMcm7。没有鉴定到Mcm8和Mcm9。同样,在苹果腐烂病菌、构巢曲霉、酵母菌中也都只鉴定到Mcm2-Mcm7,而没有鉴定到Mcm8和Mcm9。而在人、水稻、拟南芥中除了鉴定到Mcm2-Mcm7,也鉴定到Mcm8和Mcm9,这个结果与上述研究结果相一致。为了更全面了解Mcm基因家族关系,本研究将包含有真菌、植物、人的Mcm基因家族进行聚类分析,在发现的每个亚类中,葡萄座腔菌Mcm与同为真菌的苹果腐烂病菌、构巢曲霉、酵母菌Mcm关系更近。
Mcm蛋白是AAA+ATPase家族的一个亚群,形成的活性六聚体具有DNA依赖的ATP酶活性。Mcm含有一个约200个残基的高度保守的中央结构域,称之为Mcm-box。Mcm-box含有两个ATP共有的motif:Walker A [GDPXX(S/A)KS]、Walker B(IDEFDKM),在其后还有一个Mcm特有序列R-finger(SRFD)(Forsburg 2004)。本研究鉴定到的葡萄座腔菌Mcm基因家族6个成员均含有Mcm-box结构域,同时也具备Walker A、Walker B和R-finger的3个特征motif,说明鉴定到的葡萄座腔菌Mcm基因家族是正确无误的。通过分析软件预测发现,葡萄座腔菌Mcm基因家族成员均主要定位于细胞核,同时也有部分定位于高尔基体和细胞质膜。这是因为DNA复制的主要场所是细胞核,但在拟核、线粒体、叶绿体和细胞质基质(如质粒)中也可以进行DNA复制(Demesure et al. 1995),这说明葡萄座腔菌Mcm基因家族的定位与其功能相吻合。
研究表明,为了保证遗传信息在传递过程中的准确性,DNA复制需要受到严格精确的时空调控。在G1期,DNA复制解旋酶核心组分Mcm2-7六聚体在Cdc6和Cdt1的协助下被招募到复制起始识别复合体(ORC),与双链DNA结合,形成复制前复合物(pre-RC),获得复制许可资格(Bleichert et al. 2017)。进入S期,在周期蛋白依赖性激酶(Cdks)和Dbf4/Cdc7(DDK)的作用下,Cdc45和GINS复合体被招募到Mcm2-7,形成11个亚基的解旋酶全酶Cdc45-Mcm(2-7)-GINS(CMG)复合体(Costa et al. 2013;Juríková et al. 2016;Fei & Xu 2018)。随后,DNA聚合酶等其他复制相关蛋白依次被招募到CMG,共同组装成复制机器,起始DNA双向复制。因此Mcm2-7六聚体是DNA复制最重要的因子之一,该基因家族的表达会影响到细胞周期的正常进程。本研究通过qRT-PCR分析发现硫化物二甲基三硫醚处理后,葡萄座腔菌Mcm基因家族6个成员表达量下降了10.74%-73.83%。在此基础上,对葡萄座腔菌细胞周期进行检测发现,硫化物在G0/G1期对正常细胞周期发生了阻滞作用。由此可以推断,硫化物可能通过抑制葡萄座腔菌Mcm基因家族的表达,影响了Mcm2-7六聚体的正常功能,从而阻滞了正常的细胞周期进程,进而抑制了正常的细胞分裂,最终抑制了该菌的生长,并降低了病害的发生。
参考文献
Mechanisms for initiating cellular DNA replication
The complex: unwinding the mechanism of a replicative helicase
DOI:10.1128/MMBR.00019-09 URL [本文引用: 1]
Overexpression of Mcm6 predicts poor survival in patients with glioma
DOI:10.1016/j.humpath.2018.04.024 URL [本文引用: 1]
Continuous multigenerational sports selection and its mechanism reveals the mystery of ‘red fuji’ in China’s apple industry
Mechanisms for initiating cellular DNA replication
DOI:10.1146/annurev-biochem-052610-094414 URL [本文引用: 1]
A set of universal primers for amplification of polymorphic non-coding regions of mitochondrial and chloroplast DNA in plants
Role of Mcm2-7 protein phosphorylation in human cancer cells
Eukaryotic Mcm proteins: beyond replication initiation
DOI:10.1128/MMBR.68.1.109-131.2004 URL [本文引用: 3]
The role of Mcm10 in eukaryote replication origin
Sequencing of Aspergillus nidulans and comparative analysis with A. fumigatus and A. oryzae
DOI:10.1038/nature04341 URL [本文引用: 1]
Investigations on the occurrence and chemical control of Botryosphaeria canker of apple in China
Evaluation of resistance to apple ring rot in malus germplasms
Initial sequencing and analysis of the human genome
DOI:10.1038/35057062 URL [本文引用: 1]
Ki67, pcna, and Mcm proteins: markers of proliferation in the diagnosis of breast cancer
DOI:10.1016/j.acthis.2016.05.002 URL [本文引用: 3]
Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data
DOI:10.1186/1939-8433-6-4 URL [本文引用: 1]
Minichromosome maintenance 6 complex component identified by bioinformatics analysis and experimental validation in esophageal squamous cell carcinoma
DOI:10.3892/or URL [本文引用: 1]
High-resolution mapping, characterization, and optimization of autonomously replicating sequences in yeast
DOI:10.1101/gr.144659.112
PMID:23241746
[本文引用: 1]

DNA replication origins are necessary for the duplication of genomes. In addition, plasmid-based expression systems require DNA replication origins to maintain plasmids efficiently. The yeast autonomously replicating sequence (ARS) assay has been a valuable tool in dissecting replication origin structure and function. However, the dearth of information on origins in diverse yeasts limits the availability of efficient replication origin modules to only a handful of species and restricts our understanding of origin function and evolution. To enable rapid study of origins, we have developed a sequencing-based suite of methods for comprehensively mapping and characterizing ARSs within a yeast genome. Our approach finely maps genomic inserts capable of supporting plasmid replication and uses massively parallel deep mutational scanning to define molecular determinants of ARS function with single-nucleotide resolution. In addition to providing unprecedented detail into origin structure, our data have allowed us to design short, synthetic DNA sequences that retain maximal ARS function. These methods can be readily applied to understand and modulate ARS function in diverse systems.
Rapid DNA replication origin licensing protects stem cell pluripotency
DOI:10.7554/eLife.30473 URL [本文引用: 1]
Structural maintenance of chromosomes 4 is required for leukemia stem cell maintenance in mll-af9 induced acute myeloid leukemia
The Arabidopsis Information Resource (TAIR): gene structure and function annotation
DOI:10.1093/nar/gkm965 URL [本文引用: 1]
Mcm proteins in DNA replication
The MCM proteins are essential replication initiation factors originally identified as proteins required for minichromosome maintenance in Saccharomyces cerevisiae. The best known among them are a family of six structurally related proteins, MCM2-7, which are evolutionally conserved in all eukaryotes. The MCM2-7 proteins form a hexameric complex. This complex is a key component of the prereplication complex that assembles at replication origins during early G1 phase. New evidence suggests that the MCM2-7 proteins may be involved not only in the initiation but also in the elongation of DNA replication. Orchestration of the functional interactions between the MCM2-7 proteins and other components of the prereplication complex by cell cycle-dependent protein kinases results in initiation of DNA synthesis once every cell cycle.
Minichromosome maintenance protein and cancer
Proteomic analysis and nir-ii imaging of Mcm2 protein in hepatocellular carcinoma
DOI:10.1021/acs.jproteome.8b00181 URL [本文引用: 1]
Genome sequence of Valsa canker pathogens uncovers a potential adaptation of colonization of woody bark
DOI:10.1111/nph.2015.208.issue-4 URL [本文引用: 1]
Genome assembly and annotation of Botryosphaeria dothidea sdau11-99, a latent pathogen of apple fruit ring rot in China
Open-ringed structure of the cdt1- Mcm2-7 complex as a precursor of the Mcm double hexamer
DOI:10.1038/nsmb.3374 URL [本文引用: 2]
Control effect of Chinese leek extract and its main bioactive components on apple ring rot incidence
Seasonal dynamics of Botryosphaeria dothidea infections and symptom development on apple fruits and shoots in China
DOI:10.1007/s10658-016-0935-5 URL [本文引用: 1]
持续多代芽变选种及其芽变机理揭开‘红富士’在我国苹果产业独占鳌头的谜底
中国苹果枝干轮纹病发生和防治情况



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}