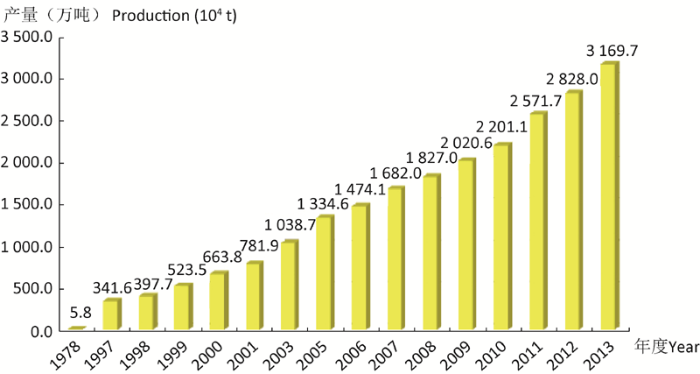
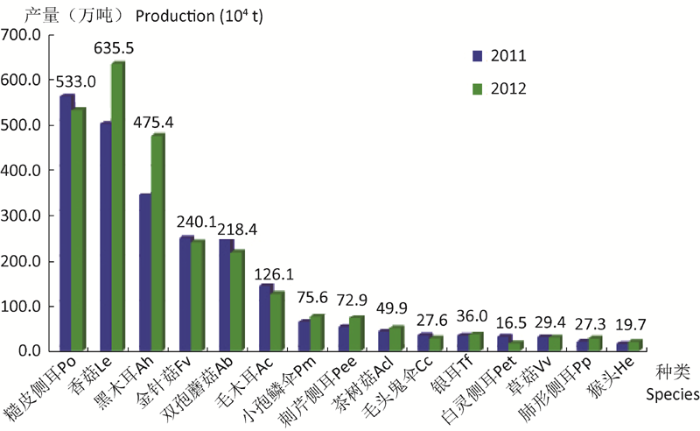
In this paper, the history of understanding and utilization of mushrooms is reviewed. China’s contribution to mushroom cultivation in early era is known to all and the development of mushroom industry at present is in progress. The development course and current situation of mushroom industry in China is presented. The basic situation of global mushroom industry and developing history, especially button mushroom industry in the West and mushroom industry in Asia is discussed. Changes in the production mode and technology of world mushroom industry are analyzed. The transfer trend of global mushroom industry from developed to developing regions will continue. China will accelerate the change of production mode into systematization, large scale, standardization and specialization.
Keywords:understanding
;
utilization
;
history
;
industry structure
;
technology transition
;
development trends
ZHANGJin-Xia, CHENQiang, HUANGChen-Yang, GAOWei, QUJi-Bin. History, current situation and trend of edible mushroom industry development[J]. 菌物学报, 2015, 34(4): 524-540 https://doi.org/10.13346/j.mycosystema.150076
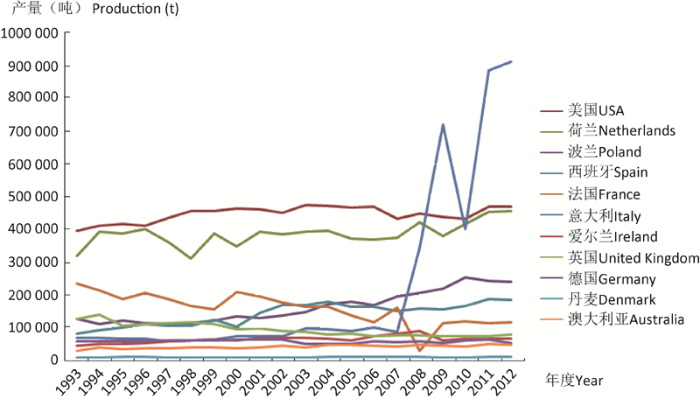
Table 2 Production of button mushroom and truffles in the West
年度 Year
产量(万吨)Production (104 t)
1938
0.4-0.5
1950
6.6
1960
13.6
1970
36
1980
87.16
1993
149.0
1994
158.5
1995
153.3
1996
157.7
1997
153.4
1998
149.9
1999
158.8
2000
159.0
2001
167.9
2002
166.6
2003
169.9
2004
173.1
2005
166.1
2006
165.0
2007
166.7
2008
188.7
2009
226.7
2010
204.0
2011
262.0
2012
263.9
Note: Resources from http://faostat3.fao.org/browse/Q/QC/E. Countries include USA, Netherlands, Poland, Spain, France, Italy, Ireland, United Kingdom, Germany, Denmark, Australia. Products include fresh and canned button mushroom and truffles.注:数据来自http://faostat3.fao.org/browse/Q/QC/E;欧美国家统计的是美国、荷兰、波兰、西班牙、法国、意大利、爱尔兰、英国、德国、丹麦、澳大利亚;产品包括鲜品、罐头和块菌.
作为大型真菌的食用菌,染色体个体小,以细胞学为基础的传统遗传学研究困难。分子生物学的发展,组学技术的不断完善,生物信息学的应用,为传统方法难以研究的食用菌的遗传学研究带来曙光。双孢蘑菇(Morin et al. 2012)、糙皮侧耳(Riley et al. 2014)、金针菇(Park et al. 2014)、草菇(Chen et al. 2013)、灵芝(Chen et al. 2012b)、香菇(未公布)、杏鲍菇(未公布)、黑木耳(未公布)、茯苓(未公布)等近10种的全基因组测序已经完成。就现有的测序数据分析,双孢蘑菇、草菇、糙皮侧耳、金针菇等食用菌的基因组规模大多在30-37Mbp,GC含量在48%-51%之间,约有10 000-13 000个预测的基因。灵芝的基因组规模略大,为43.3Mbp,GC含量为56%,预测的基因数目也更多,有16 113个。应用分子生物学技术的遗传学研究表明,草菇菌丝虽然没有锁状联合,但是仍是二极性异宗结合种类(Chen et al. 2012a)。草菇不同单孢测序表明,其交配型A因子序列的多态性显著,而B因子序列基本一致。这为草菇二极性异宗结合的性遗传特征提供了实证,颠覆了传统的同宗结合的结论。
食用菌农艺性状的遗传研究倍受业界关注。然而,在没有基因组测序之前,几乎无从开始。基因组学为这一研究奠定基础的同时,也提出了研究的突破口。根据双孢蘑菇的全基因组序列信息,发现了42个与子实体褐变黑色素合成相关的基因(Weijn et al. 2013),这些基因分布于全基因组范围内的11条染色体上;不同菌株重测序后开发出覆盖全基因组范围的SNP标记,构建了SNP标记遗传连锁图谱,在此图谱上定位了双孢蘑菇抗机械损伤的关键决定因子,为抗褐变双孢蘑菇的分子育种提供了科学依据(Gao et al. 2015)。香菇全基因组序列测定后开发出InDel分子标记,结合SRAP和TRAP标记构建了香菇的高密度遗传连锁图谱;定位出决定香菇菌丝生长速度的多个QTL位点(Gong et al. 2014),可应用于分子标记辅助育种和菌丝长速的调控研究。遗传学研究表明,糙皮侧耳的菌丝长速、酯酶同工酶位点数、子实体产量之间具显著相关性(Larraya et al. 2002),从基因组学数据中得到了进一步证实。
木质纤维素是食用菌生长发育的主要营养,多种降解酶系参与了食用菌的营养吸收与利用。研究表明:食用菌最大量的碳源来自纤维素和半纤维素。β-1,4-内切葡聚糖酶(EGL)、外切葡聚糖酶/纤维二糖水解酶(CBH)、β-葡糖苷酶(BGL)、β-1,4-木聚糖酶、β-1,4-木糖苷酶和β-甘露聚糖酶是纤维素半纤维素降解的主要酶类。然而,外包裹的木质素是纤维素和半纤维素,是食用菌降解利用的天然结构屏障。只有破坏木质素结构,才能更好地降解纤维素半纤维素。多年来,对于食用菌营养生理的研究,更多的关注在于漆酶。然而,新近污叉丝孔菌Dichomitus squalens、松生拟层孔菌Fomitopsis pinicola、灵芝Ganoderma lingzhi、桦革裥菌Lenzites betulinus、刺芹侧耳Pleurotus eryngii、糙皮侧耳Pleurotus ostreatus、云芝栓孔菌Trametes vesicolor等7种真菌对麦草中木质素的降解研究发现,污叉丝孔菌降解木质素最多,14d降解34.1%,糙皮侧耳降解木质素最少,14d降解7.1%。而降解木质素能力最差的糙皮侧耳和杏鲍菇的漆酶活性最高,其他5种真菌漆酶活性均很低,桦革裥菌甚至未检测到漆酶活性。桦革裥菌和松生拟层孔菌在培养10d时依赖型锰过氧化酶(MnP)活性最强,而糙皮侧耳MnP活性最低;污叉丝孔菌在10d时非依赖型锰过氧化酶(MnIP)活性最强,糙皮侧耳MnIP活性最低。这表明木质素的降解主要是依赖锰过氧化酶,而与漆酶关系不大(Knežević et al. 2013)。而降解的木质素最强的污叉丝孔菌本身并不能合成木质素过氧化酶(LiP),说明木质素过氧化酶也不是决定木质素降解的关键酶(Périé et al. 2008),这为食用菌营养生理的研究提供了重要的参考。
高温导致的烧菌和污染,是食用菌减产甚至绝收的重大生产问题。选育耐高温品种和创新高温伤害防控技术成为解决问题的关键。研究表明NO是白灵侧耳的高温响应信号分子,可以启动海藻糖的合成(Kong et al. 2012)。而海藻糖具缓解高温引发的氧化损伤作用(Kong et al. 2012),其以提高SOD、CAT和POD活性清除高温引发的活性氧缓解缓解氧化损伤(孟利娟等 2015)。在肺形侧耳中有TPP和TP两条海藻糖合成途径,热敏感菌株TPS活性和TPS表达显著高于耐热菌株(刘秀明等 2013),这为耐高温育种提供了必要的分子筛选标识。
Wild edible fungi - a global overview of their use and importance to people.
1
2004
... 据估测,自然界的菌物有150万种以上,其中大型真菌至少有140 000种(Hawksworth 2001).目前世界范围内存在的真菌种类约有10万种(Kirk et al. 2008),其中2 300余种为食药用菌(Boa 2004).目前我国菌物1.6万种(戴玉成和庄剑云2010),其中食用菌近1 000种(戴玉成等 2010),广泛食用的有200种左右(王向华等 2004).在远古时代,人类对食用菌的利用完全来自野生环境的采集.经历几千年对食用菌形态、生境、习性的仔细观察,人类开始了食用菌的驯化栽培,截止2004年,200种可以试验性培养,100种可以人工栽培或培养(Chang & Miles 2004),近年又增加了尖顶羊肚菌(杜习慧等 2014)和暗褐网柄牛肝菌(曹旸等 2011).实现商业化栽培的有60种左右,规模化商业栽培的有10种(Chang & Miles 2004).在报道的首次人工栽培50种中,最早的栽培记录多数在我国(表1)(Chang et al. 1993;黄年来等 1997). ...
Effect of different casing soils on the fruiting of Phlebopus portentous.
0
2011
Species clarification of the prize medicinal Ganoderma mushroom “Lingzhi”.
1
2012
... 食用菌生物多样性的研究对于我国的菌物学家是个挑战.基于Fries和Singer的大型真菌分类系统,多以形态特征为主要分类依据(Singer 1986).然而,子实体易受环境条件影响而变化的特性,给大型真菌准确的分类单元划分带来了巨大的困难.随着近代生化技术和分子生物学技术的进步,DNA特征成为新的分类依据.ITS、LSU、SSU、IGS、mtDNA、TOPⅡ、CO1、EF1α、RPB2等基因被广泛应用.新技术的应用,将香菇从Lentinus 划到了Lentinula(Pegler 1975).现代分子生物学研究表明,我国新疆阿魏Ferula sinkiangensis上发生的侧耳并非阿魏侧耳Pleurotus ferulea 单一物种,而是阿魏侧耳Pleurotus ferulea和白灵侧耳Pleurotus eryngii var. tuoliensis 2个种群(Zhang et al. 2006;赵梦然等 2015),而且后者为优势种群.研究认为,我国广泛栽培的灵芝并非欧洲的Ganoderma lucidum,而是Ganoderma lingzhi(Cao et al. 2012;戴玉成等2013);栽培多年的黑木耳也并非完全是欧洲记载的Auricularia auricula-judae,其中大部分栽培品种是独立的新种Auricularia heimuer(Wu et al. 2014;吴芳和戴玉成 2015),只有小部分品种为Auricularia auricula-judae;桑黄不是Phellinus igniarius,而是Inonotus sanghuang(戴玉成和崔宝凯 2014). ...
1
1993
... 据估测,自然界的菌物有150万种以上,其中大型真菌至少有140 000种(Hawksworth 2001).目前世界范围内存在的真菌种类约有10万种(Kirk et al. 2008),其中2 300余种为食药用菌(Boa 2004).目前我国菌物1.6万种(戴玉成和庄剑云2010),其中食用菌近1 000种(戴玉成等 2010),广泛食用的有200种左右(王向华等 2004).在远古时代,人类对食用菌的利用完全来自野生环境的采集.经历几千年对食用菌形态、生境、习性的仔细观察,人类开始了食用菌的驯化栽培,截止2004年,200种可以试验性培养,100种可以人工栽培或培养(Chang & Miles 2004),近年又增加了尖顶羊肚菌(杜习慧等 2014)和暗褐网柄牛肝菌(曹旸等 2011).实现商业化栽培的有60种左右,规模化商业栽培的有10种(Chang & Miles 2004).在报道的首次人工栽培50种中,最早的栽培记录多数在我国(表1)(Chang et al. 1993;黄年来等 1997). ...
2
2004
... 据估测,自然界的菌物有150万种以上,其中大型真菌至少有140 000种(Hawksworth 2001).目前世界范围内存在的真菌种类约有10万种(Kirk et al. 2008),其中2 300余种为食药用菌(Boa 2004).目前我国菌物1.6万种(戴玉成和庄剑云2010),其中食用菌近1 000种(戴玉成等 2010),广泛食用的有200种左右(王向华等 2004).在远古时代,人类对食用菌的利用完全来自野生环境的采集.经历几千年对食用菌形态、生境、习性的仔细观察,人类开始了食用菌的驯化栽培,截止2004年,200种可以试验性培养,100种可以人工栽培或培养(Chang & Miles 2004),近年又增加了尖顶羊肚菌(杜习慧等 2014)和暗褐网柄牛肝菌(曹旸等 2011).实现商业化栽培的有60种左右,规模化商业栽培的有10种(Chang & Miles 2004).在报道的首次人工栽培50种中,最早的栽培记录多数在我国(表1)(Chang et al. 1993;黄年来等 1997). ...
... ).实现商业化栽培的有60种左右,规模化商业栽培的有10种(Chang & Miles 2004).在报道的首次人工栽培50种中,最早的栽培记录多数在我国(表1)(Chang et al. 1993;黄年来等 1997). ...
Composition and expression of genes encoding carbohydrate-active enzymes in the straw-degrading mushroom Volvariella volvacea.
1
2013
... 作为大型真菌的食用菌,染色体个体小,以细胞学为基础的传统遗传学研究困难.分子生物学的发展,组学技术的不断完善,生物信息学的应用,为传统方法难以研究的食用菌的遗传学研究带来曙光.双孢蘑菇(Morin et al. 2012)、糙皮侧耳(Riley et al. 2014)、金针菇(Park et al. 2014)、草菇(Chen et al. 2013)、灵芝(Chen et al. 2012b)、香菇(未公布)、杏鲍菇(未公布)、黑木耳(未公布)、茯苓(未公布)等近10种的全基因组测序已经完成.就现有的测序数据分析,双孢蘑菇、草菇、糙皮侧耳、金针菇等食用菌的基因组规模大多在30-37Mbp,GC含量在48%-51%之间,约有10 000-13 000个预测的基因.灵芝的基因组规模略大,为43.3Mbp,GC含量为56%,预测的基因数目也更多,有16 113个.应用分子生物学技术的遗传学研究表明,草菇菌丝虽然没有锁状联合,但是仍是二极性异宗结合种类(Chen et al. 2012a).草菇不同单孢测序表明,其交配型A因子序列的多态性显著,而B因子序列基本一致.这为草菇二极性异宗结合的性遗传特征提供了实证,颠覆了传统的同宗结合的结论. ...
a. Sequence and comparative analysis of the MIP gene in Chinese straw mushroom, Volvariella volvacea.
1
2012
... 作为大型真菌的食用菌,染色体个体小,以细胞学为基础的传统遗传学研究困难.分子生物学的发展,组学技术的不断完善,生物信息学的应用,为传统方法难以研究的食用菌的遗传学研究带来曙光.双孢蘑菇(Morin et al. 2012)、糙皮侧耳(Riley et al. 2014)、金针菇(Park et al. 2014)、草菇(Chen et al. 2013)、灵芝(Chen et al. 2012b)、香菇(未公布)、杏鲍菇(未公布)、黑木耳(未公布)、茯苓(未公布)等近10种的全基因组测序已经完成.就现有的测序数据分析,双孢蘑菇、草菇、糙皮侧耳、金针菇等食用菌的基因组规模大多在30-37Mbp,GC含量在48%-51%之间,约有10 000-13 000个预测的基因.灵芝的基因组规模略大,为43.3Mbp,GC含量为56%,预测的基因数目也更多,有16 113个.应用分子生物学技术的遗传学研究表明,草菇菌丝虽然没有锁状联合,但是仍是二极性异宗结合种类(Chen et al. 2012a).草菇不同单孢测序表明,其交配型A因子序列的多态性显著,而B因子序列基本一致.这为草菇二极性异宗结合的性遗传特征提供了实证,颠覆了传统的同宗结合的结论. ...
b. Genome sequence of the model medicinal mushroom Ganoderma lucidum.
1
2012
... 作为大型真菌的食用菌,染色体个体小,以细胞学为基础的传统遗传学研究困难.分子生物学的发展,组学技术的不断完善,生物信息学的应用,为传统方法难以研究的食用菌的遗传学研究带来曙光.双孢蘑菇(Morin et al. 2012)、糙皮侧耳(Riley et al. 2014)、金针菇(Park et al. 2014)、草菇(Chen et al. 2013)、灵芝(Chen et al. 2012b)、香菇(未公布)、杏鲍菇(未公布)、黑木耳(未公布)、茯苓(未公布)等近10种的全基因组测序已经完成.就现有的测序数据分析,双孢蘑菇、草菇、糙皮侧耳、金针菇等食用菌的基因组规模大多在30-37Mbp,GC含量在48%-51%之间,约有10 000-13 000个预测的基因.灵芝的基因组规模略大,为43.3Mbp,GC含量为56%,预测的基因数目也更多,有16 113个.应用分子生物学技术的遗传学研究表明,草菇菌丝虽然没有锁状联合,但是仍是二极性异宗结合种类(Chen et al. 2012a).草菇不同单孢测序表明,其交配型A因子序列的多态性显著,而B因子序列基本一致.这为草菇二极性异宗结合的性遗传特征提供了实证,颠覆了传统的同宗结合的结论. ...
Notes on the nomenclature of the most widely cultivated Ganoderma species in China.
0
2013
A revised checklist of edible fungi in China.
0
2010
Numbers of fungal species hitherto known in China.
0
2010
Progress on the species of medicinal fungus Inonotus sanghuang.
0
2014
Establishment and optimization of ISSR-PCR reaction system for Auricularia polytricha.
0
2010
Quantitative trait locus mapping for bruising sensitivity and cap color of Agaricus bisporus (button mushrooms).
1
2015
... 食用菌农艺性状的遗传研究倍受业界关注.然而,在没有基因组测序之前,几乎无从开始.基因组学为这一研究奠定基础的同时,也提出了研究的突破口.根据双孢蘑菇的全基因组序列信息,发现了42个与子实体褐变黑色素合成相关的基因(Weijn et al. 2013),这些基因分布于全基因组范围内的11条染色体上;不同菌株重测序后开发出覆盖全基因组范围的SNP标记,构建了SNP标记遗传连锁图谱,在此图谱上定位了双孢蘑菇抗机械损伤的关键决定因子,为抗褐变双孢蘑菇的分子育种提供了科学依据(Gao et al. 2015).香菇全基因组序列测定后开发出InDel分子标记,结合SRAP和TRAP标记构建了香菇的高密度遗传连锁图谱;定位出决定香菇菌丝生长速度的多个QTL位点(Gong et al. 2014),可应用于分子标记辅助育种和菌丝长速的调控研究.遗传学研究表明,糙皮侧耳的菌丝长速、酯酶同工酶位点数、子实体产量之间具显著相关性(Larraya et al. 2002),从基因组学数据中得到了进一步证实. ...
Constructing a new integrated genetic linkage map and mapping quantitative trait loci for vegetative mycelium growth rate in Lentinula edodes.
1
2014
... 食用菌农艺性状的遗传研究倍受业界关注.然而,在没有基因组测序之前,几乎无从开始.基因组学为这一研究奠定基础的同时,也提出了研究的突破口.根据双孢蘑菇的全基因组序列信息,发现了42个与子实体褐变黑色素合成相关的基因(Weijn et al. 2013),这些基因分布于全基因组范围内的11条染色体上;不同菌株重测序后开发出覆盖全基因组范围的SNP标记,构建了SNP标记遗传连锁图谱,在此图谱上定位了双孢蘑菇抗机械损伤的关键决定因子,为抗褐变双孢蘑菇的分子育种提供了科学依据(Gao et al. 2015).香菇全基因组序列测定后开发出InDel分子标记,结合SRAP和TRAP标记构建了香菇的高密度遗传连锁图谱;定位出决定香菇菌丝生长速度的多个QTL位点(Gong et al. 2014),可应用于分子标记辅助育种和菌丝长速的调控研究.遗传学研究表明,糙皮侧耳的菌丝长速、酯酶同工酶位点数、子实体产量之间具显著相关性(Larraya et al. 2002),从基因组学数据中得到了进一步证实. ...
Mushrooms: the extent of the unexplored potenitial.
1
2001
... 据估测,自然界的菌物有150万种以上,其中大型真菌至少有140 000种(Hawksworth 2001).目前世界范围内存在的真菌种类约有10万种(Kirk et al. 2008),其中2 300余种为食药用菌(Boa 2004).目前我国菌物1.6万种(戴玉成和庄剑云2010),其中食用菌近1 000种(戴玉成等 2010),广泛食用的有200种左右(王向华等 2004).在远古时代,人类对食用菌的利用完全来自野生环境的采集.经历几千年对食用菌形态、生境、习性的仔细观察,人类开始了食用菌的驯化栽培,截止2004年,200种可以试验性培养,100种可以人工栽培或培养(Chang & Miles 2004),近年又增加了尖顶羊肚菌(杜习慧等 2014)和暗褐网柄牛肝菌(曹旸等 2011).实现商业化栽培的有60种左右,规模化商业栽培的有10种(Chang & Miles 2004).在报道的首次人工栽培50种中,最早的栽培记录多数在我国(表1)(Chang et al. 1993;黄年来等 1997). ...
1
1997
... 据估测,自然界的菌物有150万种以上,其中大型真菌至少有140 000种(Hawksworth 2001).目前世界范围内存在的真菌种类约有10万种(Kirk et al. 2008),其中2 300余种为食药用菌(Boa 2004).目前我国菌物1.6万种(戴玉成和庄剑云2010),其中食用菌近1 000种(戴玉成等 2010),广泛食用的有200种左右(王向华等 2004).在远古时代,人类对食用菌的利用完全来自野生环境的采集.经历几千年对食用菌形态、生境、习性的仔细观察,人类开始了食用菌的驯化栽培,截止2004年,200种可以试验性培养,100种可以人工栽培或培养(Chang & Miles 2004),近年又增加了尖顶羊肚菌(杜习慧等 2014)和暗褐网柄牛肝菌(曹旸等 2011).实现商业化栽培的有60种左右,规模化商业栽培的有10种(Chang & Miles 2004).在报道的首次人工栽培50种中,最早的栽培记录多数在我国(表1)(Chang et al. 1993;黄年来等 1997). ...
0
2010
Ainsworth & Bisby’s dictionary of the fungi. 10th edition.
1
2008
... 据估测,自然界的菌物有150万种以上,其中大型真菌至少有140 000种(Hawksworth 2001).目前世界范围内存在的真菌种类约有10万种(Kirk et al. 2008),其中2 300余种为食药用菌(Boa 2004).目前我国菌物1.6万种(戴玉成和庄剑云2010),其中食用菌近1 000种(戴玉成等 2010),广泛食用的有200种左右(王向华等 2004).在远古时代,人类对食用菌的利用完全来自野生环境的采集.经历几千年对食用菌形态、生境、习性的仔细观察,人类开始了食用菌的驯化栽培,截止2004年,200种可以试验性培养,100种可以人工栽培或培养(Chang & Miles 2004),近年又增加了尖顶羊肚菌(杜习慧等 2014)和暗褐网柄牛肝菌(曹旸等 2011).实现商业化栽培的有60种左右,规模化商业栽培的有10种(Chang & Miles 2004).在报道的首次人工栽培50种中,最早的栽培记录多数在我国(表1)(Chang et al. 1993;黄年来等 1997). ...
Lignin degradation by selected fungal species.
1
2013
... 木质纤维素是食用菌生长发育的主要营养,多种降解酶系参与了食用菌的营养吸收与利用.研究表明:食用菌最大量的碳源来自纤维素和半纤维素.β-1,4-内切葡聚糖酶(EGL)、外切葡聚糖酶/纤维二糖水解酶(CBH)、β-葡糖苷酶(BGL)、β-1,4-木聚糖酶、β-1,4-木糖苷酶和β-甘露聚糖酶是纤维素半纤维素降解的主要酶类.然而,外包裹的木质素是纤维素和半纤维素,是食用菌降解利用的天然结构屏障.只有破坏木质素结构,才能更好地降解纤维素半纤维素.多年来,对于食用菌营养生理的研究,更多的关注在于漆酶.然而,新近污叉丝孔菌Dichomitus squalens、松生拟层孔菌Fomitopsis pinicola、灵芝Ganoderma lingzhi、桦革裥菌Lenzites betulinus、刺芹侧耳Pleurotus eryngii、糙皮侧耳Pleurotus ostreatus、云芝栓孔菌Trametes vesicolor等7种真菌对麦草中木质素的降解研究发现,污叉丝孔菌降解木质素最多,14d降解34.1%,糙皮侧耳降解木质素最少,14d降解7.1%.而降解木质素能力最差的糙皮侧耳和杏鲍菇的漆酶活性最高,其他5种真菌漆酶活性均很低,桦革裥菌甚至未检测到漆酶活性.桦革裥菌和松生拟层孔菌在培养10d时依赖型锰过氧化酶(MnP)活性最强,而糙皮侧耳MnP活性最低;污叉丝孔菌在10d时非依赖型锰过氧化酶(MnIP)活性最强,糙皮侧耳MnIP活性最低.这表明木质素的降解主要是依赖锰过氧化酶,而与漆酶关系不大(Knežević et al. 2013).而降解的木质素最强的污叉丝孔菌本身并不能合成木质素过氧化酶(LiP),说明木质素过氧化酶也不是决定木质素降解的关键酶(Périé et al. 2008),这为食用菌营养生理的研究提供了重要的参考. ...
Nitric oxide is involved in the regulation of trehalose accumulation under heat stress in Pleurotus eryngii var. tuoliensis.
2
2012
... 高温导致的烧菌和污染,是食用菌减产甚至绝收的重大生产问题.选育耐高温品种和创新高温伤害防控技术成为解决问题的关键.研究表明NO是白灵侧耳的高温响应信号分子,可以启动海藻糖的合成(Kong et al. 2012).而海藻糖具缓解高温引发的氧化损伤作用(Kong et al. 2012),其以提高SOD、CAT和POD活性清除高温引发的活性氧缓解缓解氧化损伤(孟利娟等 2015).在肺形侧耳中有TPP和TP两条海藻糖合成途径,热敏感菌株TPS活性和TPS表达显著高于耐热菌株(刘秀明等 2013),这为耐高温育种提供了必要的分子筛选标识. ...
... ).而海藻糖具缓解高温引发的氧化损伤作用(Kong et al. 2012),其以提高SOD、CAT和POD活性清除高温引发的活性氧缓解缓解氧化损伤(孟利娟等 2015).在肺形侧耳中有TPP和TP两条海藻糖合成途径,热敏感菌株TPS活性和TPS表达显著高于耐热菌株(刘秀明等 2013),这为耐高温育种提供了必要的分子筛选标识. ...
Quantitative trait loci controlling vegetative growth rate in the edible basidiomycete Pleurotus ostreatus.
1
2002
... 食用菌农艺性状的遗传研究倍受业界关注.然而,在没有基因组测序之前,几乎无从开始.基因组学为这一研究奠定基础的同时,也提出了研究的突破口.根据双孢蘑菇的全基因组序列信息,发现了42个与子实体褐变黑色素合成相关的基因(Weijn et al. 2013),这些基因分布于全基因组范围内的11条染色体上;不同菌株重测序后开发出覆盖全基因组范围的SNP标记,构建了SNP标记遗传连锁图谱,在此图谱上定位了双孢蘑菇抗机械损伤的关键决定因子,为抗褐变双孢蘑菇的分子育种提供了科学依据(Gao et al. 2015).香菇全基因组序列测定后开发出InDel分子标记,结合SRAP和TRAP标记构建了香菇的高密度遗传连锁图谱;定位出决定香菇菌丝生长速度的多个QTL位点(Gong et al. 2014),可应用于分子标记辅助育种和菌丝长速的调控研究.遗传学研究表明,糙皮侧耳的菌丝长速、酯酶同工酶位点数、子实体产量之间具显著相关性(Larraya et al. 2002),从基因组学数据中得到了进一步证实. ...
A method for establishing core collection of Pleurotus ostreatus cultivated in China based on SSR markers.
0
2012
Analysis of the mating type factors in natural populations of Pleurotus eryngii var. tuoliensis in China.
0
2013
Study on the metabolic pathway of trehalose in Pleurotus pulmonarius during heat stress recovery.
1
2013
... 高温导致的烧菌和污染,是食用菌减产甚至绝收的重大生产问题.选育耐高温品种和创新高温伤害防控技术成为解决问题的关键.研究表明NO是白灵侧耳的高温响应信号分子,可以启动海藻糖的合成(Kong et al. 2012).而海藻糖具缓解高温引发的氧化损伤作用(Kong et al. 2012),其以提高SOD、CAT和POD活性清除高温引发的活性氧缓解缓解氧化损伤(孟利娟等 2015).在肺形侧耳中有TPP和TP两条海藻糖合成途径,热敏感菌株TPS活性和TPS表达显著高于耐热菌株(刘秀明等 2013),这为耐高温育种提供了必要的分子筛选标识. ...
Review and prospect of edible fungi germplasm resources in China.
0
2000
Biochemical pathway analysis of exogenous nitric oxide improving heat-tolerance of Pleurotus eryngii var. tuoliensis.
1
2015
... 高温导致的烧菌和污染,是食用菌减产甚至绝收的重大生产问题.选育耐高温品种和创新高温伤害防控技术成为解决问题的关键.研究表明NO是白灵侧耳的高温响应信号分子,可以启动海藻糖的合成(Kong et al. 2012).而海藻糖具缓解高温引发的氧化损伤作用(Kong et al. 2012),其以提高SOD、CAT和POD活性清除高温引发的活性氧缓解缓解氧化损伤(孟利娟等 2015).在肺形侧耳中有TPP和TP两条海藻糖合成途径,热敏感菌株TPS活性和TPS表达显著高于耐热菌株(刘秀明等 2013),这为耐高温育种提供了必要的分子筛选标识. ...
Genome sequence of the button mushroom Agaricus bisporus reveals mechanisms governing adaptation to a humic-rich ecological niche.
1
2012
... 作为大型真菌的食用菌,染色体个体小,以细胞学为基础的传统遗传学研究困难.分子生物学的发展,组学技术的不断完善,生物信息学的应用,为传统方法难以研究的食用菌的遗传学研究带来曙光.双孢蘑菇(Morin et al. 2012)、糙皮侧耳(Riley et al. 2014)、金针菇(Park et al. 2014)、草菇(Chen et al. 2013)、灵芝(Chen et al. 2012b)、香菇(未公布)、杏鲍菇(未公布)、黑木耳(未公布)、茯苓(未公布)等近10种的全基因组测序已经完成.就现有的测序数据分析,双孢蘑菇、草菇、糙皮侧耳、金针菇等食用菌的基因组规模大多在30-37Mbp,GC含量在48%-51%之间,约有10 000-13 000个预测的基因.灵芝的基因组规模略大,为43.3Mbp,GC含量为56%,预测的基因数目也更多,有16 113个.应用分子生物学技术的遗传学研究表明,草菇菌丝虽然没有锁状联合,但是仍是二极性异宗结合种类(Chen et al. 2012a).草菇不同单孢测序表明,其交配型A因子序列的多态性显著,而B因子序列基本一致.这为草菇二极性异宗结合的性遗传特征提供了实证,颠覆了传统的同宗结合的结论. ...
Whole genome and global gene expression analyses of the model mushroom Flammulina velutipes reveal a high capacity for lignocellulose degradation.
1
2014
... 作为大型真菌的食用菌,染色体个体小,以细胞学为基础的传统遗传学研究困难.分子生物学的发展,组学技术的不断完善,生物信息学的应用,为传统方法难以研究的食用菌的遗传学研究带来曙光.双孢蘑菇(Morin et al. 2012)、糙皮侧耳(Riley et al. 2014)、金针菇(Park et al. 2014)、草菇(Chen et al. 2013)、灵芝(Chen et al. 2012b)、香菇(未公布)、杏鲍菇(未公布)、黑木耳(未公布)、茯苓(未公布)等近10种的全基因组测序已经完成.就现有的测序数据分析,双孢蘑菇、草菇、糙皮侧耳、金针菇等食用菌的基因组规模大多在30-37Mbp,GC含量在48%-51%之间,约有10 000-13 000个预测的基因.灵芝的基因组规模略大,为43.3Mbp,GC含量为56%,预测的基因数目也更多,有16 113个.应用分子生物学技术的遗传学研究表明,草菇菌丝虽然没有锁状联合,但是仍是二极性异宗结合种类(Chen et al. 2012a).草菇不同单孢测序表明,其交配型A因子序列的多态性显著,而B因子序列基本一致.这为草菇二极性异宗结合的性遗传特征提供了实证,颠覆了传统的同宗结合的结论. ...
The classification of the genus Lentinus Fr. (Basidiomycota).
1
1975
... 食用菌生物多样性的研究对于我国的菌物学家是个挑战.基于Fries和Singer的大型真菌分类系统,多以形态特征为主要分类依据(Singer 1986).然而,子实体易受环境条件影响而变化的特性,给大型真菌准确的分类单元划分带来了巨大的困难.随着近代生化技术和分子生物学技术的进步,DNA特征成为新的分类依据.ITS、LSU、SSU、IGS、mtDNA、TOPⅡ、CO1、EF1α、RPB2等基因被广泛应用.新技术的应用,将香菇从Lentinus 划到了Lentinula(Pegler 1975).现代分子生物学研究表明,我国新疆阿魏Ferula sinkiangensis上发生的侧耳并非阿魏侧耳Pleurotus ferulea 单一物种,而是阿魏侧耳Pleurotus ferulea和白灵侧耳Pleurotus eryngii var. tuoliensis 2个种群(Zhang et al. 2006;赵梦然等 2015),而且后者为优势种群.研究认为,我国广泛栽培的灵芝并非欧洲的Ganoderma lucidum,而是Ganoderma lingzhi(Cao et al. 2012;戴玉成等2013);栽培多年的黑木耳也并非完全是欧洲记载的Auricularia auricula-judae,其中大部分栽培品种是独立的新种Auricularia heimuer(Wu et al. 2014;吴芳和戴玉成 2015),只有小部分品种为Auricularia auricula-judae;桑黄不是Phellinus igniarius,而是Inonotus sanghuang(戴玉成和崔宝凯 2014). ...
Purification and characterization of laccases from the white-rot basidiomyceteDichomitus squalens
1
1998
... 木质纤维素是食用菌生长发育的主要营养,多种降解酶系参与了食用菌的营养吸收与利用.研究表明:食用菌最大量的碳源来自纤维素和半纤维素.β-1,4-内切葡聚糖酶(EGL)、外切葡聚糖酶/纤维二糖水解酶(CBH)、β-葡糖苷酶(BGL)、β-1,4-木聚糖酶、β-1,4-木糖苷酶和β-甘露聚糖酶是纤维素半纤维素降解的主要酶类.然而,外包裹的木质素是纤维素和半纤维素,是食用菌降解利用的天然结构屏障.只有破坏木质素结构,才能更好地降解纤维素半纤维素.多年来,对于食用菌营养生理的研究,更多的关注在于漆酶.然而,新近污叉丝孔菌Dichomitus squalens、松生拟层孔菌Fomitopsis pinicola、灵芝Ganoderma lingzhi、桦革裥菌Lenzites betulinus、刺芹侧耳Pleurotus eryngii、糙皮侧耳Pleurotus ostreatus、云芝栓孔菌Trametes vesicolor等7种真菌对麦草中木质素的降解研究发现,污叉丝孔菌降解木质素最多,14d降解34.1%,糙皮侧耳降解木质素最少,14d降解7.1%.而降解木质素能力最差的糙皮侧耳和杏鲍菇的漆酶活性最高,其他5种真菌漆酶活性均很低,桦革裥菌甚至未检测到漆酶活性.桦革裥菌和松生拟层孔菌在培养10d时依赖型锰过氧化酶(MnP)活性最强,而糙皮侧耳MnP活性最低;污叉丝孔菌在10d时非依赖型锰过氧化酶(MnIP)活性最强,糙皮侧耳MnIP活性最低.这表明木质素的降解主要是依赖锰过氧化酶,而与漆酶关系不大(Knežević et al. 2013).而降解的木质素最强的污叉丝孔菌本身并不能合成木质素过氧化酶(LiP),说明木质素过氧化酶也不是决定木质素降解的关键酶(Périé et al. 2008),这为食用菌营养生理的研究提供了重要的参考. ...
Extensive sampling of basidiomycete genomes demonstrates inadequacy of the white-rot/brown-rot paradigm for wood decay fungi.
1
2014
... 作为大型真菌的食用菌,染色体个体小,以细胞学为基础的传统遗传学研究困难.分子生物学的发展,组学技术的不断完善,生物信息学的应用,为传统方法难以研究的食用菌的遗传学研究带来曙光.双孢蘑菇(Morin et al. 2012)、糙皮侧耳(Riley et al. 2014)、金针菇(Park et al. 2014)、草菇(Chen et al. 2013)、灵芝(Chen et al. 2012b)、香菇(未公布)、杏鲍菇(未公布)、黑木耳(未公布)、茯苓(未公布)等近10种的全基因组测序已经完成.就现有的测序数据分析,双孢蘑菇、草菇、糙皮侧耳、金针菇等食用菌的基因组规模大多在30-37Mbp,GC含量在48%-51%之间,约有10 000-13 000个预测的基因.灵芝的基因组规模略大,为43.3Mbp,GC含量为56%,预测的基因数目也更多,有16 113个.应用分子生物学技术的遗传学研究表明,草菇菌丝虽然没有锁状联合,但是仍是二极性异宗结合种类(Chen et al. 2012a).草菇不同单孢测序表明,其交配型A因子序列的多态性显著,而B因子序列基本一致.这为草菇二极性异宗结合的性遗传特征提供了实证,颠覆了传统的同宗结合的结论. ...
The Agaricales in modern taxonomy
1
1986
... 食用菌生物多样性的研究对于我国的菌物学家是个挑战.基于Fries和Singer的大型真菌分类系统,多以形态特征为主要分类依据(Singer 1986).然而,子实体易受环境条件影响而变化的特性,给大型真菌准确的分类单元划分带来了巨大的困难.随着近代生化技术和分子生物学技术的进步,DNA特征成为新的分类依据.ITS、LSU、SSU、IGS、mtDNA、TOPⅡ、CO1、EF1α、RPB2等基因被广泛应用.新技术的应用,将香菇从Lentinus 划到了Lentinula(Pegler 1975).现代分子生物学研究表明,我国新疆阿魏Ferula sinkiangensis上发生的侧耳并非阿魏侧耳Pleurotus ferulea 单一物种,而是阿魏侧耳Pleurotus ferulea和白灵侧耳Pleurotus eryngii var. tuoliensis 2个种群(Zhang et al. 2006;赵梦然等 2015),而且后者为优势种群.研究认为,我国广泛栽培的灵芝并非欧洲的Ganoderma lucidum,而是Ganoderma lingzhi(Cao et al. 2012;戴玉成等2013);栽培多年的黑木耳也并非完全是欧洲记载的Auricularia auricula-judae,其中大部分栽培品种是独立的新种Auricularia heimuer(Wu et al. 2014;吴芳和戴玉成 2015),只有小部分品种为Auricularia auricula-judae;桑黄不是Phellinus igniarius,而是Inonotus sanghuang(戴玉成和崔宝凯 2014). ...
SNP analysis of the fragments of three functional genes of the mushroom Lentinula edodes.
0
2013
Color atlas of wild commercial mushrooms in Yunnan.
0
2004
Melanin biosynthesis pathway in Agaricus bisporus mushrooms.
1
2013
... 食用菌农艺性状的遗传研究倍受业界关注.然而,在没有基因组测序之前,几乎无从开始.基因组学为这一研究奠定基础的同时,也提出了研究的突破口.根据双孢蘑菇的全基因组序列信息,发现了42个与子实体褐变黑色素合成相关的基因(Weijn et al. 2013),这些基因分布于全基因组范围内的11条染色体上;不同菌株重测序后开发出覆盖全基因组范围的SNP标记,构建了SNP标记遗传连锁图谱,在此图谱上定位了双孢蘑菇抗机械损伤的关键决定因子,为抗褐变双孢蘑菇的分子育种提供了科学依据(Gao et al. 2015).香菇全基因组序列测定后开发出InDel分子标记,结合SRAP和TRAP标记构建了香菇的高密度遗传连锁图谱;定位出决定香菇菌丝生长速度的多个QTL位点(Gong et al. 2014),可应用于分子标记辅助育种和菌丝长速的调控研究.遗传学研究表明,糙皮侧耳的菌丝长速、酯酶同工酶位点数、子实体产量之间具显著相关性(Larraya et al. 2002),从基因组学数据中得到了进一步证实. ...
Notes on the nomenclature of the Auricularia auricula-judae complex.
1
2015
... 食用菌生物多样性的研究对于我国的菌物学家是个挑战.基于Fries和Singer的大型真菌分类系统,多以形态特征为主要分类依据(Singer 1986).然而,子实体易受环境条件影响而变化的特性,给大型真菌准确的分类单元划分带来了巨大的困难.随着近代生化技术和分子生物学技术的进步,DNA特征成为新的分类依据.ITS、LSU、SSU、IGS、mtDNA、TOPⅡ、CO1、EF1α、RPB2等基因被广泛应用.新技术的应用,将香菇从Lentinus 划到了Lentinula(Pegler 1975).现代分子生物学研究表明,我国新疆阿魏Ferula sinkiangensis上发生的侧耳并非阿魏侧耳Pleurotus ferulea 单一物种,而是阿魏侧耳Pleurotus ferulea和白灵侧耳Pleurotus eryngii var. tuoliensis 2个种群(Zhang et al. 2006;赵梦然等 2015),而且后者为优势种群.研究认为,我国广泛栽培的灵芝并非欧洲的Ganoderma lucidum,而是Ganoderma lingzhi(Cao et al. 2012;戴玉成等2013);栽培多年的黑木耳也并非完全是欧洲记载的Auricularia auricula-judae,其中大部分栽培品种是独立的新种Auricularia heimuer(Wu et al. 2014;吴芳和戴玉成 2015),只有小部分品种为Auricularia auricula-judae;桑黄不是Phellinus igniarius,而是Inonotus sanghuang(戴玉成和崔宝凯 2014). ...
Species clarification of the most important and cultivated Auricularia mushroom “Heimuer”: evidence from morphological and molecular data.
1
2014
... 食用菌生物多样性的研究对于我国的菌物学家是个挑战.基于Fries和Singer的大型真菌分类系统,多以形态特征为主要分类依据(Singer 1986).然而,子实体易受环境条件影响而变化的特性,给大型真菌准确的分类单元划分带来了巨大的困难.随着近代生化技术和分子生物学技术的进步,DNA特征成为新的分类依据.ITS、LSU、SSU、IGS、mtDNA、TOPⅡ、CO1、EF1α、RPB2等基因被广泛应用.新技术的应用,将香菇从Lentinus 划到了Lentinula(Pegler 1975).现代分子生物学研究表明,我国新疆阿魏Ferula sinkiangensis上发生的侧耳并非阿魏侧耳Pleurotus ferulea 单一物种,而是阿魏侧耳Pleurotus ferulea和白灵侧耳Pleurotus eryngii var. tuoliensis 2个种群(Zhang et al. 2006;赵梦然等 2015),而且后者为优势种群.研究认为,我国广泛栽培的灵芝并非欧洲的Ganoderma lucidum,而是Ganoderma lingzhi(Cao et al. 2012;戴玉成等2013);栽培多年的黑木耳也并非完全是欧洲记载的Auricularia auricula-judae,其中大部分栽培品种是独立的新种Auricularia heimuer(Wu et al. 2014;吴芳和戴玉成 2015),只有小部分品种为Auricularia auricula-judae;桑黄不是Phellinus igniarius,而是Inonotus sanghuang(戴玉成和崔宝凯 2014). ...
0
2004
0
2002
Making SSR fingerprint profile for commercial cultivars of Lentinula edodes.
0
2012
0
2010
0
1999
0
2009
Genetic polymorphism of ferula mushroom growing on Ferula sinkiangensis.
1
2006
... 食用菌生物多样性的研究对于我国的菌物学家是个挑战.基于Fries和Singer的大型真菌分类系统,多以形态特征为主要分类依据(Singer 1986).然而,子实体易受环境条件影响而变化的特性,给大型真菌准确的分类单元划分带来了巨大的困难.随着近代生化技术和分子生物学技术的进步,DNA特征成为新的分类依据.ITS、LSU、SSU、IGS、mtDNA、TOPⅡ、CO1、EF1α、RPB2等基因被广泛应用.新技术的应用,将香菇从Lentinus 划到了Lentinula(Pegler 1975).现代分子生物学研究表明,我国新疆阿魏Ferula sinkiangensis上发生的侧耳并非阿魏侧耳Pleurotus ferulea 单一物种,而是阿魏侧耳Pleurotus ferulea和白灵侧耳Pleurotus eryngii var. tuoliensis 2个种群(Zhang et al. 2006;赵梦然等 2015),而且后者为优势种群.研究认为,我国广泛栽培的灵芝并非欧洲的Ganoderma lucidum,而是Ganoderma lingzhi(Cao et al. 2012;戴玉成等2013);栽培多年的黑木耳也并非完全是欧洲记载的Auricularia auricula-judae,其中大部分栽培品种是独立的新种Auricularia heimuer(Wu et al. 2014;吴芳和戴玉成 2015),只有小部分品种为Auricularia auricula-judae;桑黄不是Phellinus igniarius,而是Inonotus sanghuang(戴玉成和崔宝凯 2014). ...
Comparison studies of genetic diversity of Pleurotus eryngii var. tuoliensis by IGS2-RFLP, SCoT and ISSR markers.
0
2013
Genetic diversity analysis of wild ferula mushroom based on IGS2-RFLP markers and cultural characteristics.
0
2015
不同覆土的性质及其对暗褐网柄牛肝菌人工栽培出菇的影响
1
2011
... 据估测,自然界的菌物有150万种以上,其中大型真菌至少有140 000种(Hawksworth 2001).目前世界范围内存在的真菌种类约有10万种(Kirk et al. 2008),其中2 300余种为食药用菌(Boa 2004).目前我国菌物1.6万种(戴玉成和庄剑云2010),其中食用菌近1 000种(戴玉成等 2010),广泛食用的有200种左右(王向华等 2004).在远古时代,人类对食用菌的利用完全来自野生环境的采集.经历几千年对食用菌形态、生境、习性的仔细观察,人类开始了食用菌的驯化栽培,截止2004年,200种可以试验性培养,100种可以人工栽培或培养(Chang & Miles 2004),近年又增加了尖顶羊肚菌(杜习慧等 2014)和暗褐网柄牛肝菌(曹旸等 2011).实现商业化栽培的有60种左右,规模化商业栽培的有10种(Chang & Miles 2004).在报道的首次人工栽培50种中,最早的栽培记录多数在我国(表1)(Chang et al. 1993;黄年来等 1997). ...
中国灵芝学名之管见
1
2013
... 食用菌生物多样性的研究对于我国的菌物学家是个挑战.基于Fries和Singer的大型真菌分类系统,多以形态特征为主要分类依据(Singer 1986).然而,子实体易受环境条件影响而变化的特性,给大型真菌准确的分类单元划分带来了巨大的困难.随着近代生化技术和分子生物学技术的进步,DNA特征成为新的分类依据.ITS、LSU、SSU、IGS、mtDNA、TOPⅡ、CO1、EF1α、RPB2等基因被广泛应用.新技术的应用,将香菇从Lentinus 划到了Lentinula(Pegler 1975).现代分子生物学研究表明,我国新疆阿魏Ferula sinkiangensis上发生的侧耳并非阿魏侧耳Pleurotus ferulea 单一物种,而是阿魏侧耳Pleurotus ferulea和白灵侧耳Pleurotus eryngii var. tuoliensis 2个种群(Zhang et al. 2006;赵梦然等 2015),而且后者为优势种群.研究认为,我国广泛栽培的灵芝并非欧洲的Ganoderma lucidum,而是Ganoderma lingzhi(Cao et al. 2012;戴玉成等2013);栽培多年的黑木耳也并非完全是欧洲记载的Auricularia auricula-judae,其中大部分栽培品种是独立的新种Auricularia heimuer(Wu et al. 2014;吴芳和戴玉成 2015),只有小部分品种为Auricularia auricula-judae;桑黄不是Phellinus igniarius,而是Inonotus sanghuang(戴玉成和崔宝凯 2014). ...
药用真菌桑黄种类研究
1
2014
... 食用菌生物多样性的研究对于我国的菌物学家是个挑战.基于Fries和Singer的大型真菌分类系统,多以形态特征为主要分类依据(Singer 1986).然而,子实体易受环境条件影响而变化的特性,给大型真菌准确的分类单元划分带来了巨大的困难.随着近代生化技术和分子生物学技术的进步,DNA特征成为新的分类依据.ITS、LSU、SSU、IGS、mtDNA、TOPⅡ、CO1、EF1α、RPB2等基因被广泛应用.新技术的应用,将香菇从Lentinus 划到了Lentinula(Pegler 1975).现代分子生物学研究表明,我国新疆阿魏Ferula sinkiangensis上发生的侧耳并非阿魏侧耳Pleurotus ferulea 单一物种,而是阿魏侧耳Pleurotus ferulea和白灵侧耳Pleurotus eryngii var. tuoliensis 2个种群(Zhang et al. 2006;赵梦然等 2015),而且后者为优势种群.研究认为,我国广泛栽培的灵芝并非欧洲的Ganoderma lucidum,而是Ganoderma lingzhi(Cao et al. 2012;戴玉成等2013);栽培多年的黑木耳也并非完全是欧洲记载的Auricularia auricula-judae,其中大部分栽培品种是独立的新种Auricularia heimuer(Wu et al. 2014;吴芳和戴玉成 2015),只有小部分品种为Auricularia auricula-judae;桑黄不是Phellinus igniarius,而是Inonotus sanghuang(戴玉成和崔宝凯 2014). ...
中国食用菌名录
0
2010
中国菌物已知种数
1
2010
... 据估测,自然界的菌物有150万种以上,其中大型真菌至少有140 000种(Hawksworth 2001).目前世界范围内存在的真菌种类约有10万种(Kirk et al. 2008),其中2 300余种为食药用菌(Boa 2004).目前我国菌物1.6万种(戴玉成和庄剑云2010),其中食用菌近1 000种(戴玉成等 2010),广泛食用的有200种左右(王向华等 2004).在远古时代,人类对食用菌的利用完全来自野生环境的采集.经历几千年对食用菌形态、生境、习性的仔细观察,人类开始了食用菌的驯化栽培,截止2004年,200种可以试验性培养,100种可以人工栽培或培养(Chang & Miles 2004),近年又增加了尖顶羊肚菌(杜习慧等 2014)和暗褐网柄牛肝菌(曹旸等 2011).实现商业化栽培的有60种左右,规模化商业栽培的有10种(Chang & Miles 2004).在报道的首次人工栽培50种中,最早的栽培记录多数在我国(表1)(Chang et al. 1993;黄年来等 1997). ...

 , 陈强, 黄晨阳, 高巍, 曲积彬
, 陈强, 黄晨阳, 高巍, 曲积彬



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}